Эта статья обеспечивает надежный анализ, чтобы сообщить об относительной экспрессии на клеточной поверхности мембранных белков, таких как ионные каналы, выраженные в рекомбинантных клетках с использованием существующей технологии проточной цитометрии. Ионные каналы являются пороформирующие мембранные белки, которые отвечают за регулирование электрических сигналов с помощью стробирования поток ионов через клеточную мембрану. Они классифицируются по механизму активации, природы и селективности видов ионов, проходящих через поры, где они локализованы. На клеточном и тканевом уровнях, макроскопические потоки ионов через ионные каналы являются продуктом биофизической (стробирования и проникания), биохимические (фосфорилирования), и биогенеза (синтез, гликозилирование, торговля людьми и деградации) свойств 1. Каждый из этих процессов является уникальным для каждого типа ионных каналов и оптимизирована для выполнения физиологической роли ионного канала. Следовательно, изменения в любой из этих доработаны процессов черезунаследованные или генные модификации, часто упоминается как "channelopathy", может быть вредным для клеточного гомеостаза. Важно подчеркнуть, что обеспечение «правильного» количества ионных каналов на поверхности клетки имеет решающее значение для клеточного гомеостаза. Даже небольшое увеличение (коэффициента усиления из-функции) и небольшое снижение (с потерей функции) в канале активности ионов имеют потенциал, чтобы вызвать серьезные патологии в течение жизни. Дефекты в доставке на клеточной поверхности зрелых ионных каналов является важным фактором , определяющим в многочисленных каналопатии, таких как муковисцидоз (CFTR ионного канала) 2 и сердечных аритмий длинного интервала QT формы синдрома (сердечных калиевых каналов) 3.
Каналопатии связаны с сердечной внезапной смерти 4. В настоящее время во всем мире распространенность всех сердечных каналопатии считается по меньшей мере , 1: 2,000-1: 3000 в индивидуальной 5 и несут ответственность за примерно половину внезапной сердечной смерти от аритмии окSES 6. Дисфункция сердечного напряжения закрытого калиевые, натрий-и кальциевые селективные ионные каналы, как известно, играют ключевую роль в этом процессе. 1.2 потенциалзависимыми кальциевых каналов L-типа Ca V необходим для инициации синхронизированного сокращения мышц сердца. Сердечный L-типа Ca V 1,2 канал белковый комплекс мульти-субъединица состоит из основного порообразующего Са V & alpha ; 1 субъединицы и Ca V ß и Ca V α2δ1 вспомогательных подразделений 7-12. Обратите внимание , что полный комплект вспомогательных подразделений требуется для производства функциональных Ca V 1,2 каналов в плазматической мембране и динамические взаимодействия между этими субъединицами имеют важное значение для поддержания нормальной электрической функции сердца 13. Ca V ß способствует поверхности экспрессии клеток Са V 1,2 каналов через нековалентному наномолярной гидрофобного взаимодействия 14. Co-выражение V α2δ1 субъединиц Wi Caго Са V SS переплете Са V α1 стимулирует экспрессию пикового тока ( от 5 до 10 раз) и способствует активации канала при более отрицательных напряжений. Усиление-оф-функции мутации порообразующего субъединицей Ca V 1.2 были связаны с формой желудочковой аритмией называется длинный QT синдром 15 в то время как множество точечных мутаций в трех основных субъединиц , образующих L-типа Ca V 1.2 канала были выявлены у пациентов , страдающих от аритмий короткого интервала QT формы синдрома 16,17. Ионные каналы являются мембранные белки, которые могут быть исследованы с биохимической перспективной (белковой химии) с использованием или электрофизиологических инструментов (вольт-генераторные машины) и часто используя эти дополнительные подходы. Электрофизиологии, в частности , цельноклеточной патч-зажим, является подходящим подход к выяснению функции ионных каналов 15 , но не может разрешить изменения в области незаконного оборота белка с изменениями в их биофизическихсвойства. химии белков, однако, часто ограниченное применение из-за относительно низкой экспрессии больших мембранных белков относительно небольших растворимых белков. Надежные методы с высокой пропускной способностью с помощью флуоресцентной считывания необходимо разработать для того, чтобы конкретно рассмотреть дефекты белка биогенезе вызывая изменения в клеточной поверхности экспрессии ионных каналов.
Проточная цитометрия является биофизической технологии , используемой в подсчета клеток, сортировки, обнаружения биомаркеров и белковой инженерии 18. Когда раствор образца живых клеток или частиц вводят в проточном цитометре, клетки упорядочены в единый поток , который может быть зондируется системой обнаружения машины (рис 1). Первый поток цитометра прибора , произведенного в 1956 году 19 обнаружен только один параметр , но современные проточные цитометры несколько лазеров и детекторов флуоресценции , которые позволяют обнаруживать более 30 флуоресцентных параметров 20,21.Фильтры и зеркала (эмиссионные оптика) направляют рассеяния света или лампы дневного света клеток к электронной сети (фотодиод и детекторы), которые преобразуют свет пропорционально его интенсивности. Цифровые данные анализируются с помощью специализированного программного обеспечения и первичный выход отображается как точка участка 21.
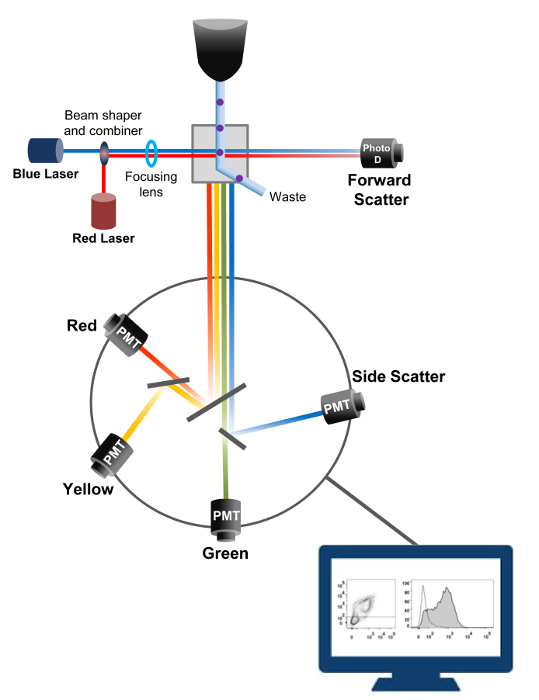
Рисунок 1:. Биофизические принципы проточной цитометрии сортировки Отдельные клетки выталкиваются через сопло под высоким давлением в потоке оболочки текучей среды , которая перемещает их через один или несколько лазерных точек допроса. Световой пучок отклоняется проходящими клеток и света, собираемого в прямом направлении (вперед Scatter, FCS), отправляется на фотодиод, который преобразует свет в сигнал, пропорциональный размеру клетки. Свет также собирается под углом к лазерному пути 90 ° и направлен детекторы (называемые также умножители (ФЭУ)).Этот свет проходит через дихроичных зеркал, которые позволяют обнаружение бокового рассеяния сигнала (SSC), который отражает детализацию в клетках, а также флуоресцентных излучений, если возбужденные флуорохромы присутствуют в клетке. Три детекторы (зеленый, желтый и красный) представлены с разными фильтрами длины волны полосовых, что позволяет одновременное обнаружение различных флюорохромами. Различные сигналы преобразуются в цифровую форму с помощью внешнего компьютера и преобразуются в данные , которые будут проанализированы для количественного определения характеристик ячеек. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
Высокой пропускная способность цитометров была использована для количественного определения относительной экспрессии мембранного рекомбинантного дикого типа и торговли людьми с дефицитом напряжения закрытого L-типа Ca V 1.2 каналов и связанных с ними субъединиц в живых клетках. кДНК создает совместноеDing для белков были двукратно помеченный одновременно нести внеклеточный нефлуоресцентного эпитоп, который может быть обнаружен с помощью непроницаемого флуоресцентного конъюгированных антител и внутриклеточный флуорофором, который конститутивно флуоресцентный. Оба внеклеточный эпитоп, вставленный во внеклеточной петле белка, и внутриклеточный флуорофор, вставленный после С-конца, пересчитаны с белком. В этой серии экспериментов V α2δ1 белок Са сконструированные для экспрессии внеклеточного гемагглютинина (HA) эпитоп (YPYDVPDYA) обнаружен непроницаемым FITC (флуоресцеин изотиоцианатом) -conjugated анти-HA и mCherry как внутренняя внутриклеточного флуорофора. Для определения относительного уровня экспрессии на клеточной поверхности клиновидного α2δ1 mCherry-Ca HA-меченый белок, рекомбинантные клетки , экспрессирующие слитый белок , собирали после трансфекции, и окрашивали ФИТЦ-конъюгированного мышиных моноклональных анти-HA эпитоп тег antibodY (рисунок 2). FITC представляет собой органическое соединение, которое флуоресцентный значительно меньше, чем ферментных репортеров и, следовательно, как вероятно, не препятствовать биологической функции. mCherry- Ca V α2δ1-HA избыточно экспрессируется в TSA-201cells, производит значительное увеличение 3-журнала в флуоресценции FITC и mCherry флуоресценции на двумерных участков 22. Учитывая, что эпитоп HA находится во внеклеточной части белка, интенсивность флуоресценции для FITC, полученный в присутствии неповрежденных клеток отражают относительный показатель клеточной поверхностной экспрессии HA-меченого белка. Доступность эпитопа HA в конструкциях систематически проверяется путем измерения сигнала FITC после клеточной проницаемости. Эта мера также служит для подтверждения нормализованное общее экспрессию белка, так как относительные интенсивности флуоресценции для FITC оценивается в проницаемыми клеток качественно сопоставимы с относительными значениями флуоресценции Foг mCherry измеряется при проницаемыми и непермеабилизированных условиях 22,23. Важно отметить, что собственный спектр флуоресценции смещается в сторону более высоких значений после того, как пермеабилизации но сообщается единственное значение является изменение интенсивности флуоресценции по сравнению с конструкцией управления. Относительные изменения интенсивности флуоресценции для испытаний конструкций оцениваются с использованием интенсивности флуоресценции (ΔMean ΔMFI) значения для каждого флуорофора (mCherry или FITC). Эксперименты предназначены для измерения интенсивности флуоресценции тест конструкта по отношению к интенсивности флуоресценции конструкции управления выраженной в тех же условиях, чтобы ограничить экспериментальные вариации в собственной флуоресценции флуорофора-конъюгированных антител. Два мембранных белков были успешно изучены с помощью этого анализа: в качестве порообразующего субъединицей напряжения закрытого кальциевых каналов L-типа Ca V 1,2 14,22 и в другой серииЭксперименты, внеклеточный Ca вспомогательный V α2δ1 субъединица 22,23. Следующий протокол использовали для определения экспрессии клеточной поверхности Са V α2δ1 субъединицей сердца L-типа Ca V 1,2 каналов в контрольных условиях и после мутаций , влияющих на посттрансляционной модификации ионного канала. В рамках стандартных экспериментальных условиях клеточной поверхности флуоресценции FITC увеличивает квази- линейно с выражением кДНК , кодирующей mCherry-Ca V α2δ1-HA белков (рисунок 5 из ссылки 22).
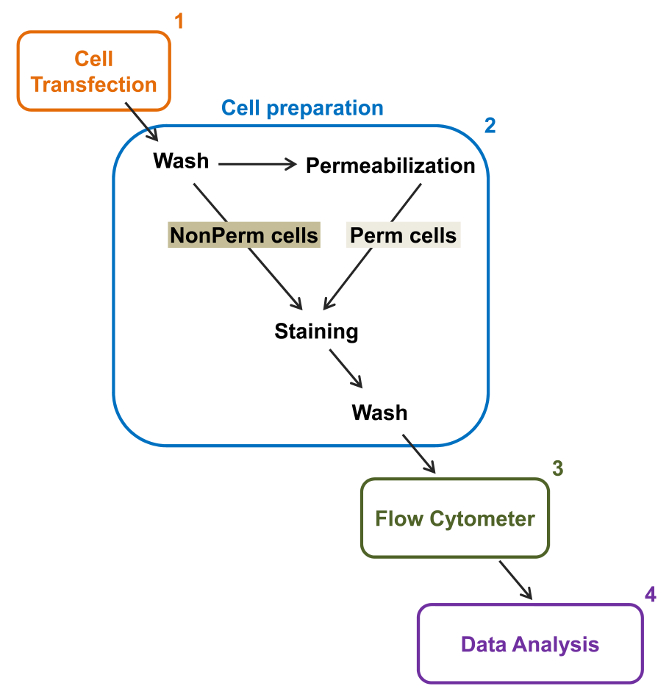
Рис . 2: Схематическое представление полной и мембранной маркировки в проточной цитометрии экспериментальный протокол Схема описывает некоторые из основных шагов , необходимых для количественной оценки относительной общей и поверхностной клеточной экспрессии рекомбинантных ионных каналов Ф.Л.вл цитометрии. Клетки трансфицируют с двойным меченого строительство mCherry-Ca V α2δ1-HA в TSA-201 клеток (1) и окрашивают до или после того, как пермеабилизации (2). Многопараметрические данные получают в проточном цитометре (3) для многомерного анализа (4). Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
