Ce document fournit un test fiable pour signaler l'expression relative de la surface cellulaire des protéines membranaires telles que des canaux ioniques exprimés dans les cellules recombinantes en utilisant la technologie de cytométrie de flux existant. Les canaux ioniques sont des protéines membranaires porogènes qui sont responsables de la commande des signaux électriques par gating le flux d'ions à travers la membrane cellulaire. Ils sont classés par le mécanisme d'activation, la nature et la sélectivité des espèces d'ions qui transitent à travers le pore où ils sont localisés. Au niveau cellulaire et tissulaire, les flux ioniques macroscopiques à travers les canaux ioniques sont le produit de biophysique propriétés 1 (gating et perméation), biochimique (phosphorylation), et la biogenèse (synthèse, glycosylation, le trafic et la dégradation). Chacun de ces processus est unique pour tous les types de canaux ioniques et elle est optimisée pour remplir le rôle physiologique du canal ionique. Par conséquent, des modifications dans l'un de ces processus affiné grâce à unhéréditaire ou une modification génétique, souvent appelée "canalopathie", peut être préjudiciable à l'homéostasie cellulaire. Il est important de souligner que la prestation du «droit» quantité de canaux ioniques à la surface cellulaire est essentielle pour l'homéostasie cellulaire. Même de petites augmentations (gain de fonction) et de petites diminutions (perte de fonction) dans l'activité des canaux ioniques ont le potentiel de provoquer une pathologie grave au cours d'une vie. Défauts dans la livraison de la surface cellulaire des canaux ioniques matures est un déterminant important dans de nombreux channelopathies, comme la fibrose kystique (canal ionique CFTR) 2 et les arythmies cardiaques de la forme longue de syndrome du QT (canaux potassiques cardiaques) 3.
Channelopathies sont associés à la mort subite cardiaque 4. La prévalence mondiale actuelle de tous les channelopathies cardiaques est considéré comme au moins 1: 2,000-1: 3.000 par personne 5 et sont responsables d'environ la moitié d' un coup arythmique cardiaque mort caSES 6. Dysfonction cardiaque en voltage-dépendants de sodium, de potassium et de calcium canaux ioniques sélectifs sont connus pour jouer un rôle clé dans ce processus. Le 1.2 canaux calciques voltage-dépendants de type L Ca V est nécessaire pour engager synchronisé coeur la contraction musculaire. L cardiaque de type Ca 1,2 V de canal est un complexe protéique sous-unités multiples composée de la sous – unité principale Ca porogène V α1 et Ca Vss et Ca V α2δ1 sous – unités auxiliaires 7-12. Notez que la liste complète des sous – unités auxiliaires est nécessaire pour produire fonctionnels Ca V 1.2 canaux à la membrane plasmique et les interactions dynamiques entre ces sous – unités sont essentiels pour soutenir la fonction électrique normale du cœur 13. Ca Vss favorise l'expression de surface cellulaire de Ca 1,2 V canaux à travers une nanomolaire interaction hydrophobe non covalente 14. Co-expression de la Ca V sous – unité α2δ1 wiCa th Ca V ß lié V α1 stimule l' expression du courant de crête (5 à 10 fois) et favorise l' activation du canal à des tensions négatives. Gain de fonction des mutations de la sous – unité de formation de pores Ca V 1.2 ont été associés à une forme d'arythmie ventriculaire appelé le syndrome du QT long 15 tandis qu'une foule de mutations ponctuelles dans les trois principales sous – unités formant le type L Ca V 1.2 canaux ont été identifiés chez des sujets souffrant d'arythmies du QT court formulaire de syndrome 16,17. Les canaux ioniques sont des protéines membranaires qui peuvent être étudiés du point de vue biochimique (chimie des protéines) ou à l'aide d'outils électrophysiologiques (machines génératrices de courant) et souvent l'aide de ces approches complémentaires. Électrophysiologie, notamment la cellule entière patch-serrage, est une approche appropriée pour élucider la fonction des canaux ioniques 15 mais ne peut pas résoudre des modifications dans le trafic de protéines à partir des changements dans leur biophysiquePropriétés. la chimie des protéines a cependant souvent un usage limité en raison de la relativement faible expression de grandes protéines de la membrane par rapport aux protéines solubles plus petits. méthodes à haut débit robustes en utilisant la fluorescence de lecture doivent être développées afin de traiter spécifiquement les défauts de la protéine biogenèse provoquant des changements dans l'expression de la surface cellulaire des canaux ioniques.
La cytométrie en flux est une technologie biophysique employée dans le comptage des cellules, le tri, la détection de biomarqueurs et l' ingénierie des protéines 18. Lorsqu'une solution de cellules vivantes ou des particules de l' échantillon est injecté dans un cytomètre de flux, les cellules sont ordonnées en un flux unique qui peut être sondé par le système de détection de la machine (figure 1). Le premier instrument cytomètre produit en 1956 19 un seul paramètre détecté , mais cytomètres de flux modernes ont de multiples lasers et les détecteurs de fluorescence permettant la détection de plus de 30 paramètres fluorescents 20,21.Filtres et miroirs (optique d'émission) dirigent la diffusion de la lumière ou de la lumière fluorescente de cellules à un réseau électronique (photodiode et détecteurs) qui convertissent la lumière proportionnellement à son intensité. Les données numériques sont analysées à l' aide de logiciels spécialisés et la sortie primaire est affiché comme un complot point 21.
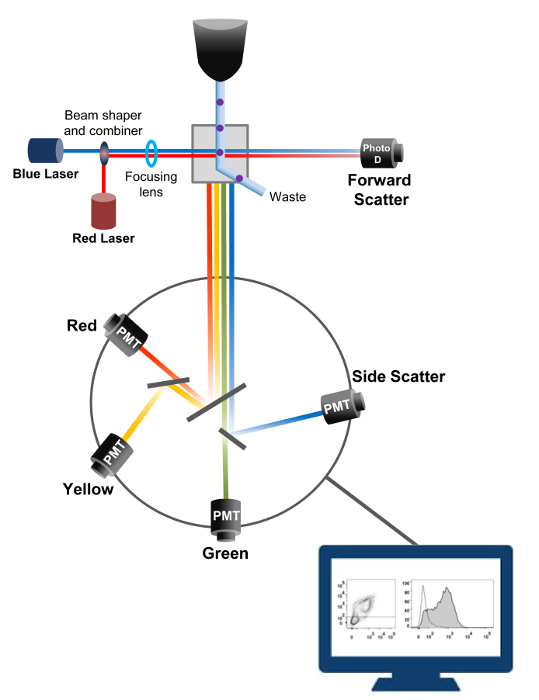
Figure 1:. Principes biophysiques des flux de tri cytométrie Les cellules individuelles sont poussées à travers une buse sous haute pression dans un courant de fluide de gaine qui les déplace à travers un ou plusieurs points d'interrogation laser. Le faisceau lumineux est dévié par les cellules qui passent et la lumière collectée dans le sens avant (Forward Scatter, FCS) est envoyé à une photodiode qui convertit la lumière en un signal proportionnel à la taille de la cellule. La lumière est également recueillie à un angle de la trajectoire du laser 90 ° et envoyé aux détecteurs (également appelés photomultiplicateurs (PMT)).Cette lumière est acheminée à travers des miroirs dichroïques qui permettent la détection du signal de diffusion latérale (SSC), qui reflète la granularité dans les cellules, et les émissions fluorescentes si fluorochromes excités sont présents dans la cellule. Trois détecteurs (vert, jaune et rouge) sont représentés avec différents filtres de longueur d'onde de bande passante, ce qui permet la détection simultanée de différents fluorochromes. Les différents signaux sont numérisés par un ordinateur externe et converties en données qui seront analysées afin de quantifier les caractéristiques des cellules. S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
La capacité à haut débit de cytomètres de flux a été exploité pour quantifier l'expression membranaire relative de type sauvage recombinant et de type L voltage-dépendants trafic déficient Ca V 1.2 canaux et sous – unités associées à des cellules vivantes. constructions d'ADNc de coding pour les protéines ont été doublement marquées pour transporter simultanément un épitope extracellulaire non fluorescent qui peut être détecté par un anticorps conjugué fluorescent imperméable et un fluorophore intracellulaire qui est constitutivement fluorescente. À la fois l'épitope extracellulaire, inséré dans une boucle extracellulaire de la protéine, et le fluorophore intracellulaire, après avoir inséré l'extrémité C-terminale, sont convertis avec la protéine. Dans cette série d'expériences, la protéine V α2δ1 Ca a été conçu pour exprimer une hémagglutinine extracellulaire (HA) épitope (YPYDVPDYA) détectée par un matériau imperméable au FITC (isothiocyanate de fluorescéine) conjugué à anti-HA et mCherry comme fluorophore intrinsèque intracellulaire. Pour déterminer le niveau d'expression de surface cellulaire relative du V α2δ1 mCherry-Ca protéine HA étiquetée, des cellules recombinées exprimant la protéine de fusion ont été récoltées après la transfection, et colorées avec l'anticorps monoclonal de souris anti-HA étiquette d' épitope Antibod conjugué au FITCy (Figure 2). FITC est un composé organique fluorescent qui est considérablement plus petite que les journalistes enzymes et donc moins susceptibles d'interférer avec la fonction biologique. mCherry- Ca V α2δ1-HA surexprimé dans Tsa-201cells, produit une augmentation de 3 log significative de la fluorescence FITC et mCherry fluorescence sur des parcelles en deux dimensions 22. Etant donné que l'épitope HA est situé dans la partie extracellulaire de la protéine, l'intensité de fluorescence FITC obtenue en présence de cellules intactes reflètent l'indice relatif de l'expression de surface cellulaire de la protéine HA étiquetée. L'accessibilité de l'épitope HA dans les constructions est systématiquement validée en mesurant le signal FITC après perméabilisation cellulaire. Cette mesure permet aussi de confirmer l'expression de la protéine totale normalisée depuis les intensités de fluorescence relatives pour FITC estimées dans les cellules perméabilisées sont qualitativement comparables aux valeurs de fluorescence relative for mCherry mesurée sous perméabilisées et non perméabilisées conditions 22,23. Il est important de noter que le spectre de fluorescence intrinsèque est déplacée vers des valeurs plus élevées après perméabilisation, mais que la seule valeur étant rapportée est la variation de l'intensité de la fluorescence par comparaison avec la construction témoin. Les changements relatifs à l'intensité de fluorescence pour les constructions d'essai sont estimées en utilisant l'intensité ΔMean Fluorescence (de ΔMFI) des valeurs pour chaque fluorophore (mCherry ou FITC). Les expériences sont conçues pour mesurer l'intensité de la fluorescence du produit d'assemblage de test par rapport à l'intensité de fluorescence du produit d'assemblage de commande exprimée dans les mêmes conditions expérimentales pour limiter les variations de la fluorescence intrinsèque de l'anticorps conjugué à un fluorophore. Deux protéines de la membrane ont été étudiés avec succès en utilisant ce dosage: la sous – unité formant des pores du canal calcium de type L voltage-dépendants Ca 1,2 V 14,22 et dans une autre série d'expériences, l'auxiliaire extracellulaire Ca V α2δ1 sous – unité 22,23. Le protocole suivant a été utilisé pour déterminer l'expression de surface cellulaire de la sous – unité V α2δ1 Ca du type L cardiaque Ca V 1.2 canal dans des conditions de contrôle et d' après des mutations affectant la modification post – traductionnelle du canal ionique. Dans des conditions expérimentales normalisées, la fluorescence à la surface cellulaire de FITC augmente quasi linéairement avec l'expression de l' ADNc codant pour les mCherry-Ca V protéines α2δ1-HA (figure 5 de la référence 22).
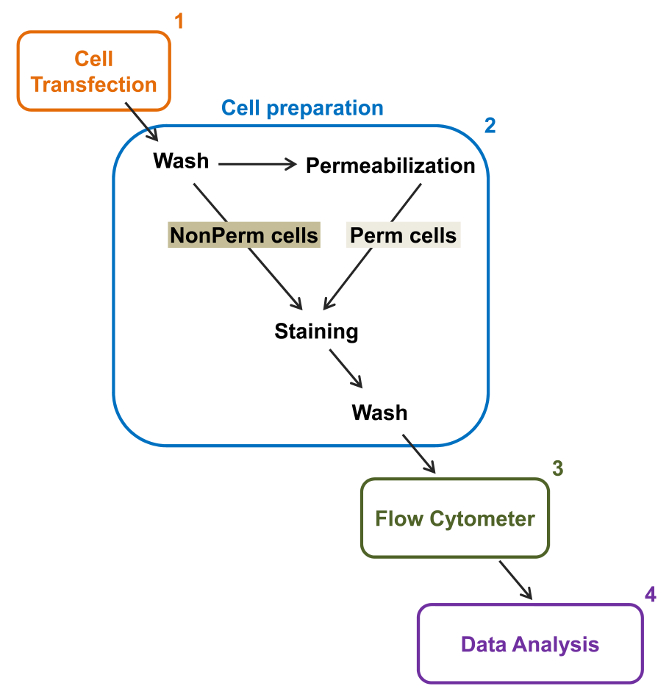
Figure 2:. Représentation schématique de l' étiquetage totale et de la membrane dans la cytométrie protocole expérimental Le schéma décrit quelques – unes des principales mesures nécessaires pour quantifier l'expression relative totale et surface cellulaire des canaux ioniques recombinantes par flow cytométrie. Les cellules sont transfectées avec la construction double marquage mCherry-Ca V α2δ1-HA dans Tsa-201 cellules (1) et colorées avant ou après perméabilisation (2). Les données multiparamètres sont acquises dans un cytomètre de flux (3) pour l' analyse multivariée (4). S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
