Wachstumsanalyse hängt von einer Reihe von Werkzeugen, die von Pflanzenwissenschaftler häufig verwendet werden, Genotyp bestimmt Wachstumsunterschiede und / oder phänotypische Reaktionen auf Umweltfaktoren zu beschreiben. Dazu gehören Größe und Gewichtsmessungen der gesamten Anlage oder ein Organ und Berechnungen der Wachstumsraten, die zugrunde liegenden Mechanismen des Wachstums zu erkunden. Organwachstum wird durch Zellteilung und Expansion auf der zellulären Ebene bestimmt. Daher analysiert die Quantifizierung dieser beiden Prozesse in Wachstum einschließlich ist der Schlüssel zum Verständnis der Unterschiede in Vollorganwachstum 1. Folglich ist es wichtig, eine geeignete Methodik haben zellulären Wachstumsparameter zu bestimmen, der relativ leicht von nicht spezialisierten Labors zu verwenden.
Kinematic Analyse wurde bereits als Ansatz etabliert 2 einen leistungsstarken Rahmen für die Entwicklung von Organwachstumsmodelle bieten. Die Technik wurde für lineare Systeme optimiert,wie Arabidopsis thaliana Wurzeln und monokotyle Blätter, sondern auch für nichtlineare Systeme, wie dikotyle Blätter 3. Heute wird diese Methode immer mehr, wie genetische Studie verwendet wird, hormonelle, entwicklungs und Umweltfaktoren beeinflussen die Zellteilung und die Expansion in verschiedenen Organen (Tabelle 1). Darüber hinaus bietet sie auch einen Rahmen zelluläre Prozesse auf die ihnen zugrunde liegenden biochemischen, molekularen und physiologischen Bestimmungen (Tabelle 2) zu verbinden, obwohl Einschränkungen können durch Organgröße und räumliche Organisation für Techniken , die höhere Mengen an Pflanzenmaterial erfordern auferlegt werden (zB Metaboliten Messungen, Proteomics, etc.).
Monokotylen Blätter, wie beispielsweise Mais (Zea mays) Blatt-, für lineare Systeme , in denen Zellen von der Basis des Blattes gegen die Spitze zu bewegen, der Reihe nach durch die Meristem und Dehnungszone Leiten des reifen zu erreichenZone. Dies macht es zu einem idealen Modellsystem für quantitative Untersuchungen der räumlichen Muster des Wachstums 4. Darüber hinaus haben Maisblätter große Wachstumszonen (Meristem und Dehnungszone über mehrere Zentimeter 5) und in anderen Organisationsebenen Möglichkeiten für Studien. Dies ermöglicht die Untersuchung der (vermeintlichen) Regulationsmechanismen der Zellteilung steuern und Expansion, durch kinematische Analyse durch eine Reihe von molekularen Techniken quantifiziert, physiologische Messungen und Zellbiologie Ansätze (Tabelle 2).
Hier stellen wir ein Protokoll für eine kinematische Analyse in monokotylen Blätter durchführt. Zuerst wird erklärt, wie eine richtige Analyse der sowohl die Zellteilung und Zell Dehnung als Funktion der Position längs der Blattachse zu leiten und wie kinematischen Parameter zu berechnen. Zweitens zeigen wir auch, wie diese als Grundlage für die Stichprobenplan verwendet werden können. Hier diskutieren wir zwei Fälle: hochauflösende Abtastung eind konzentriert Probenahme, verbesserte Dateninterpretation und die Einsparung von Zeit / Geld ermöglicht, respectively.
Tabelle 1 Übersicht der kinematischen Analysen Methoden zur Quantifizierung der Zellteilung und Expansion in verschiedenen Organen.
| Organ | Referenz |
| monokotylen Blätter | 16, 20, 21, 22 |
| Wurzelspitzen | 2, 23, 24, 25, 26, 27, 28, 29 |
| dikotylen Blätter | 21, 30, 31 |
| Sprossapikalmeristem | 32 |
Tabelle 1 Übersicht der kinematischen Analysen Methoden zur Quantifizierung der Zellteilung und Expansion in verschiedenen Organen.
<p class="jove_content" fo:keep-together.within-page = "1">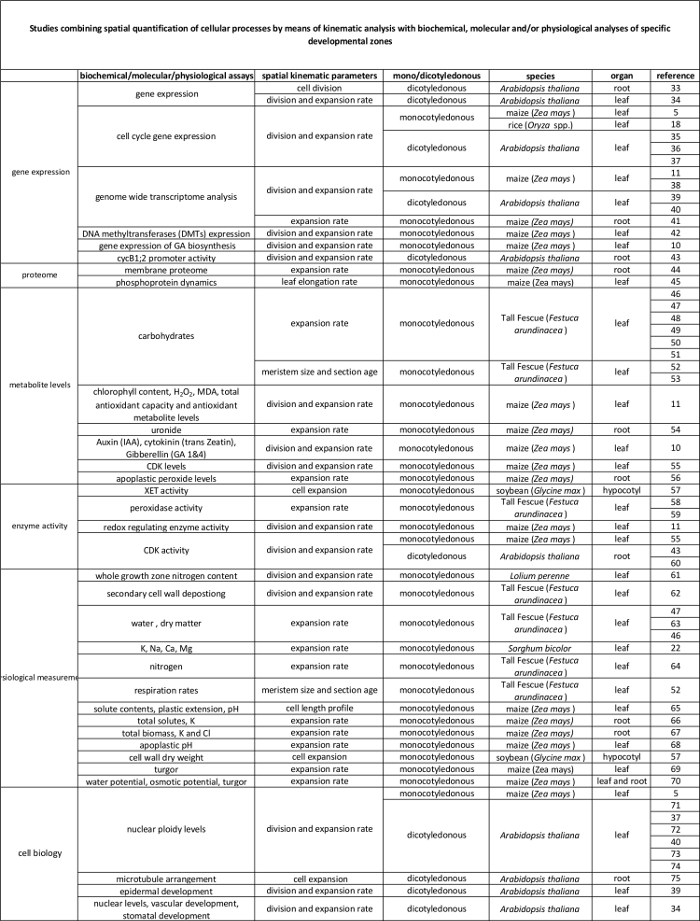
Tabelle 2 Zusammenhang zwischen zellulärer Prozesse quantifiziert durch die kinematische Analyse auf ihre Regulation auf molekularer Ebene. Verweise auf verschiedene Studien , die die Quantifizierung von zellulären Prozessen zu den Ergebnissen von biochemischen und molekularen Assays in verschiedenen Spezies und Organen. Xyloglucan endotransglucosylase (XET), Malondialdehyd (MDA), Cyclin-abhängigen Kinasen (CDK). Bitte hier klicken , um eine größere Version dieser Tabelle anzuzeigen.
