Fotosynthetische organismen zoals diatomeeën moeten omgaan met veranderende lichtomstandigheden en reageren met verfijnde acclimatisering mechanismen die fotosynthetische hoogrenderende ondersteunen en beschermen tegen foto-oxidatieve schade veroorzaakt door het overmatige licht. Een grote licht-beschermende proces in fotosynthetische eukaryoten is de hoge energie blussen (qE) van geabsorbeerde licht dat zich als de belangrijkste bijdrage aan de niet-fotochemische blussen (NPQ) onder lichte stress voorwaarden1,2 voordoet ,3. De lichte oogsten antenne complexen (LHC) zijn betrokken bij de regulering van excitatie energie overdracht trajecten. In antwoord op hoge licht geïnduceerde lage pH in de chloroplast lumen, de antenne systeem schakelopties van het licht oogsten van status naar status het blussen. Deze dissipatief energietoestand beschermt photosystems (PS) en andere complexen in het membraan van thylakoïde foto-oxidatie. In fotosynthetische eukaryoten, wordt de qE meestal veroorzaakt door twee factoren1,–2,3. Een factor is het gespecialiseerde licht oogsten eiwit dat op de lage pH reageert. De publieke omroepen proteïne induceert de qE in hogere planten4. LhcSRs5, gemoduleerd door publieke omroepen activiteit, veroorzaken de qE in groene algen6. Diatomeeën bezitten Lhcx-achtige eiwitten die structuur verwant zijn aan de LHCSRs7,,8,,9,10.
De tweede factor van qE is de Xantofyl-cyclus waar carotenoïden van de antenne door de-epoxidatie omgezet in de vorm van een foto-beschermende en teruggekeerd door epoxidatie. In planten en groene algen, wordt Violaxanthine geconverteerd naar zeaxanthine. In diatomeeën, wordt diadinoxanthin geconverteerd naar diatoxanthin, die vervolgens met de omvang van NPQ11 correleert. Het licht van de Diatomee oogsten antenne bezit enkele eigenaardigheden, hoewel het evolutionaire gerelateerde te planten en algen LHCs. De overgang van licht aan foto-bescherming is enorm snel en de capaciteit van NPQ is hoger in vergelijking met planten12. Dit zou een reden waarom diatomeeën zijn ijzersterk in verschillende ecologische niches op een manier die zij verantwoordelijk voor 45% van de oceanische netto primaire productie13 zijn. Diatomee lichte oogsten systemen zijn dus een interessant object voor fotosynthese onderzoek.
Diatomeeën, zoals de centric soorten Cyclotella meneghiniana, intrinsieke licht thylakoïde oogsten systemen vernoemd naar de pigmenten die ze bezitten bind – fucoxanthin, chlorofyl (chl) a en c, vandaar FCP. Light oogsten van eiwitten, zoals FCPs, zijn ingebed in de thylakoïde membraan systeem dat bestaat uit verschillende lagen van het membraan. Diatomeeën vormen bands van drie thylakoiden. Dit complex situatie maakt het moeilijk om ze te bestuderen op moleculair niveau in het membraan van thylakoïde. Bovendien, vele componenten dragen bij aan de verordening van licht oogsten (zie hierboven). Daarom waren de complexen in vele benaderingen, geïsoleerd van het membraan met behulp van milde wasmiddelen, bijvoorbeeld n-Dodecyl-β-D-maltopyranoside (β-DDM), die solubilize van het membraan, maar de FCP-complexen intact te houden. Vele spectroscopische studies werden uitgevoerd met behulp van ontbindend FCP te onderzoeken intramoleculaire energie overdracht14,15,16,17. Deze voormalige aanpak was echter beperkt omdat de regulering van energie-overdracht excitonic interactie met andere antenne complexen of photosystems moet. Vandaar worden dit soort studies niet uitgevoerd met ontbindend complexen omdat de interactie tussen complexen verloren is.
Een belangrijk kenmerk in antenne verordening is de “moleculaire verdringing” van de antenne en photosystems in de thylakoïde membraan18. Voorheen, een eenvoudige benadering werd uitgevoerd om te simuleren van dit effect in vitro. Het wasmiddel werd verwijderd, wat leidt tot willekeurige aggregatie van antenne complexen. Hoewel sommige redelijke gegevens werd verkregen door deze aanpak17,19, de wasmiddel verwijdering weerspiegelt niet de situatie in vivo en kent een aantal beperkingen omdat het complexen zijn niet in hun reguliere tertiaire interactie structuur.
Het gebruik van liposomen overwint verscheidene van de vroegere beperkingen. De tertiaire structuur is nog volledig intact. Het liposoom membraan biedt een quasi-eigen omgeving voor de antenne-complexen. Het membraan scheidt de binnenkant van de liposomen van de buitenomgeving. Langs deze weg bieden liposomen twee reactie compartimenten voor studies van ion en pH verlopen ook wat transportprocessen. Verder kunnen de parameters van het experimentele systeem gemakkelijker worden beheerd voor studies in het membraan van thylakoïde. Liposomen bleken al te zijn een uitstekend hulpmiddel om te studeren fotosynthetische complexen. Een belangrijk aandachtspunt in het verleden was op de plant waar het effect van de gewijzigde lipide samenstelling is getest op LHC II20LHC. In andere benaderingen waren de eiwit-eiwit interactie tussen verschillende LHC II onderzochte21. Ook werden sommige studies in groene algen uitgevoerd die beschrijven spontane clustering tussen LHC22. Gezien het belang van diatomeeën voor aquatische ecosystemen, relatief weinig studies werden uitgevoerd met antenne complexen van diatomeeën. Twee studies onderzocht de complexen van de antenne van de centric Cyclotella meneghiniana, waar de clustering van de FCP antenne23 en reactievermogen van FCP elektrochemische verlopen24 werden getoond. Dus, liposomen zijn een uitstekend hulpmiddel om te studeren Diatomee antennes en hun interactie en de verordening in bijna oorspronkelijke omstandigheden. De liposomen zijn veelzijdig sinds vele aandoeningen zoals lipide samenstelling, grootte van de liposomen, eiwit dichtheid en de omliggende waterfase kan worden gecontroleerd. De methode vereist bovendien lage bedragen van monsters. Het experimentele systeem is robuust en zeer reproduceerbaar. De opdeling van liposomen zorgt voor het bestuderen van pH en ion hellingen, die zijn belangrijke factoren in de regulering van de antenne complexen.
Hier beschrijven we de isolatie van FCP antenne complexen van C. meneghiniana en hun opneming in liposomen met natuurlijke thylakoïde lipide samenstelling. Ook wij voorbeeldige gegevens leveren voor de spectroscopische karakterisering van ontbindend FCP en vergelijk deze met FCP in liposomen. De methode geeft een overzicht van kennis en gestandaardiseerde protocollen verkregen van de verbeteringen van de Gundermann en Büchel 201223, Natali et al. 201622en Ahmad en Dietzel 201724.
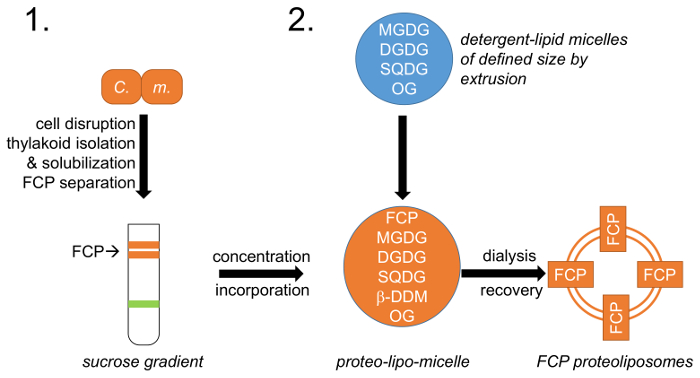
Figuur 1: schematische weergave van de werkstroom. (1) verwijst naar alinea 1 die beschrijft celgroei, verstoring en thylakoïde isolatie met FCP scheiding aanleiding sacharose dichtheid verlopen; C. m. –Cyclotella meneghiniana cellen. (2) voorbereiding van natuurlijke thylakoïde lipide mengsel (MGDG, DGDG en SQDG) als beschreven in lid 2 en oprichting van lipide-wasmiddel micellen met octylglycoside (OG). Een gedefinieerde lipide-micel grootte wordt bereikt door met behulp van membranen met een diameter van gedefinieerde porie extrusie. FCP en lipide-micellen zijn herenigd op een vooraf gedefinieerde lipide: eiwitverhouding en de detergentia OG en β-DDM zijn verwijderd via gecontroleerde dialyse FCP proteoliposomes vormen. Klik hier voor een grotere versie van dit cijfer.
