단일 분자 Förster 공명 에너지 전송 (smFRET)는 개별 분자의 수준에서 두 염료 -a 기증자와 수용자 사이의 FRET 효율을 측정하는 기술이다. FRET는 두 염료의 중복 에너지 스펙트럼에서 발생하는 광물리 학적 과정입니다 : 기증자는 특정 파장의 빛에 흥분하고 수용자에서 방출의 결과로, 수용자로 비 방사 적으로 에너지를 전송합니다. 이 전송의 효율은 두 염료 사이의 거리의 여섯 번째 힘에 반비례하므로 전송 효율은 distance1에 따라 다릅니다. 따라서, 이러한 FRET 효율은 염료가 부착되는 분자에 대한 공간 정보를 결정하는 데 사용될 수 있다. 이러한 규모와 FRET 효율의 변화가 Angstrom 분자 운동에 민감하다는 사실은, 핵산 및 단백질과 같은 생체 분자에 대한 구조적 정보를 평균 4,5,6의 합병증없이 조사하는 데 적합한 기술을 만든다. 상대적인 FRET 효율성의 변화는 생체 분자 상호 작용 및 형성 역학을 모니터링하는 데 사용될 수 있지만, 단백질(un)접기, 전사 및 DNA 복제 및 수리와 같은 주요 세포 공정에 빛을 흘리는 반면, 절대 FRET 효율은 생체 분자 구조 결정에 대한 정확한 거리를 결정하는 데 사용되어 왔습니다7,8,9,10,111 , 다른 구조적 방법에 필요한 결정화 또는 동결의 필요성을 극복4,12.
smFRET 실험은 가장 일반적으로 두 가지 형태를 취, 공초점 또는 총 내부 반사 형광 (TIRF) 현미경 검사. 두 접근 사이 생체 분자의 분자 역학은 전형적으로 pico-to millisecond (공초점, 자유롭게 확산 분자)에서 시간까지 시간 척도에서 조사 할 수 있습니다 (TIRF, 표면 고정 분자). 이는 각 기술에 관련된 다양한 설정 때문입니다. TIRF 현미경 검사법에서 분자는 슬라이드 표면에 고정되어 에반센드 파에 흥분됩니다(그림 1A). 여기서, 그러나, 초점은 이 smfBox의 형식이기 때문에 공초점 현미경 검사법에 있습니다. 공초점 현미경 검사에서 분자는 고숫자 조리개 렌즈를 통해 레이저 빔을 솔루션 내의 일부 지정된 깊이에서 스팟으로 집중하여 형성된 공초점 부피(~1fL)를 통해 브라우니아 운동을 통해 자유롭게 확산된다(그림 1B). 생성된 방출은 동일한 조리개를 통해 다시 초점을 맞추고 이색 거울을 통해 필터링됩니다(전체 회로도의 그림 1C ). 그런 다음 초점이 맞지 않는 빛을 제거하고 눈사태 포토다이오드(APD)에 초점을 맞춥니다. APD가 광자를 감지하면 TTL 펄스를 출력하며, 타이밍은 최대 피코초 해상도로 기록될 수 있습니다. 공초점 부피 부근내에서 자유롭게 확산되는 분자의 관찰 시간은 일반적으로 밀리초 의 순서 내에 있습니다.
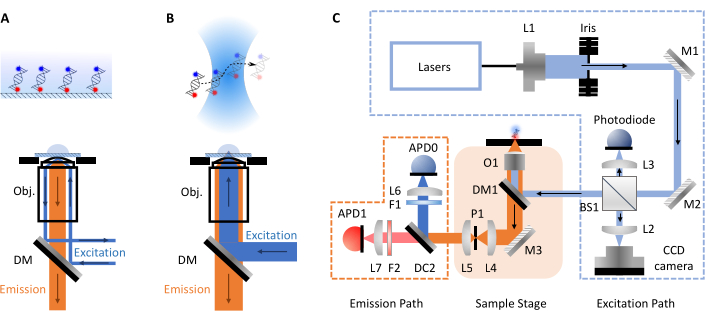
그림 1: 현미경 검사법과 smfBox 설정의 원리를 보여주는 회로도. (A) 총 내부 반사 형광 (TIRF) 현미경 원리 : 흥분 빛은 목표 (Obj.)의 가장자리로 지시하고 기하급수적으로 부패 하는 에반스 엔스 필드를 생성하는 커버 슬립 버퍼 인터페이스에서 총 내부 반사를 겪는다. (B) 공초점 현미경 검사: 분자를 자유롭게 확산시키는 것은 시료에 초점을 맞춘 거의 회절 제한 지점에 의해 흥분됩니다. (C) 눈사태 포토다이오드(APD), 빔 스플리터(BS), 이색 거울(DM), 필터(F), 미러(M), 목표(O) 및 핀홀(P)의 모든 주요 구성 요소를 보여주는 이 프로토콜에 사용되는 smfBox 설정. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
최근에는 smFRET 기술이 두 가지 색 여기를 통합하여 기증자와 수용자 내분 파장을 일치시키는 레이저가 번갈아 5입니다. 이것은 두 가지 방법 중 하나에서 수행 할 수 있습니다, KHz 기간에 연속 파 레이저를 변조하여 첫 번째, 이는 교대 레이저 여기로 알려진 (ALEX)13,14. 두 번째 방법은 MHz 시간 척도에서 빠른 펄스를 인터링합니다. 이는 나노초-ALEX15 또는 펄스 인터리브 된 여기 (PIE)16입니다. 이러한 모든 접근법에서, 수용자 레이저의 정보는 소위 stoichiometry의 계산으로 이끌어 냅니다, 이는 낮은 FRET 효율을 가진 분자와 수용자가 부족한 사람들 (불완전한 라벨링 또는 광표백을 통해)를 구별할 수 있는. PIE/ns-ALEX를 사용하면 단일 분자 수준에서 형광 수명에 추가로 접근할 수 있으며, 편광 광학과 결합될 때 이산화정도를 측정할 수 있습니다. 이러한 측정 조합은 다중 파라미터 형광 검출(MFD)9라고 합니다.
smFRET의 많은 장점에도 불구 하 고, 그것은 널리 상업 악기의 높은 비용과 간단한의 부족으로 전문 실험실 외부에서 사용 되지 않습니다., 자체 빌드 대안. 저가형 오픈소스 현미경 검사법의 개발을 향한 추세가 증가하고 있으며 플랑크톤코프17, 오픈플렉스어마이크로스코프18, 플렉시코프19, miCube20, liteTIRF21 및 Squid22를 포함한 다른 플랫폼이 최근 등장했습니다. 본 연구에서는 단일 분자를 자유롭게 확산시키는 데 대한 두 염료 사이의 FRET 효율을 측정할 수 있는 최근 개발된 비용 효율적인 공초점 설정인 smfBox를 사용하기 위한 프로토콜을 설명합니다. 자세한 빌드 지침과 필요한 모든 운영 소프트웨어는 https://craggslab.github.io/smfBox/ 23에서 자유롭게 사용할 수 있습니다. smfBox의 광학 배열은 저렴하고 널리 접근 가능한 제조업체에서 구입한 쉽게 사용할 수 있는 구성 요소에서 조립되며, 현미경 본체(표준 공초점 설정에서 비용의 대부분을 담당함)는 사용자 정의 조명 이밀한 양극산화 알루미늄 상자로 대체되었습니다(주변 조명 조건에서 측정할 수 있음). 이 상자에는 여기 이색, 목표 및 핀홀, 기계식 레이저 인터록을 포함한 주요 광학 부품이 포함되어 있어 Class I 레이저 제품으로 안전한 작동을 가능하게 합니다(전체 회로도의 그림 1C 참조). smfBox는 ALEX를 사용하여 염료 스토이치오메트리의 유효성을 검사하고 정확한 FRET 보정 요인을 결정합니다. 데이터 수집의 모든 측면을 제어하고 많은 타사 분석 도구와 호환되는 오픈 소스 광자-HDF5 format24의 데이터를 출력하는 맞춤형 오픈 소스 소프트웨어(smOTTER)를 사용하여 작동됩니다. smfBox 및 수집 및 데이터 분석 프로토콜은 최근 멀티 랩 블라인드 연구에서 >20 개의 다른 기기 (공초점 및 TIRF 모두)에 대해 테스트되었습니다25. 얻은 FRET 효율성은 smfBox가 시판되는 설정 가격의 일부만 비용에도 불구하고 다른 모든 악기와 우수한 계약을 맺었습니다.
여기서 단계별 프로토콜은 smfBox를 사용하여 DNA 듀플렉스를 자유롭게 확산시키는 데 있어 정확하고 절대적인 FRET 효율성을 획득하고 분석하기 위해 설명되어 있으며, 전원켜기부터 정렬 및 초점, 데이터 수집 및 분석에 이르기까지 모든 방법을 제공합니다. 여기서 사용되는 샘플은 세계 적인 맹목적인 연구에서 평가된 3개의 이중 DN(고, 중간 및 저FRET 효율성을 전시하고, 표 1참조)입니다; 그러나, 이 방법은 단백질 및 기타 핵산을 포함한 많은 분자 시스템에 적응할 수 있다. 희망은 이러한 상세한 프로토콜, smfBox23에 대한 기존의 기존 빌드 지침과 함께, 실험실의 넓은 범위에 더 액세스 할 수있는이 강력한 기술을 만드는 데 도움이 될 것입니다.
