Strongyloides stercoralis долгое время игнорировался как важный патоген человека по сравнению с более широко признанными анкилостомами и круглым червем Ascaris lumbricoides1. Предыдущие исследования глистной нагрузки часто сильно недооценивали распространенность S. stercoralis из-за низкой чувствительности общих методов диагностики S. stercoralis2. В последние годы эпидемиологические исследования, основанные на улучшенных диагностических инструментах, показали, что истинная распространенность инфекций S. stercoralis намного выше, чем сообщалось ранее, примерно 610 миллионов человек во всем мире2.
Как S. stercoralis, так и другие виды Strongyloides, включая близкородственного крысиного паразита и общую лабораторную модель S. ratti, имеют необычный жизненный цикл, который выгоден для экспериментальных геномных исследований, поскольку он состоит как из паразитических, так и из свободноживущих (экологических) поколений3 (рисунок 1). В частности, как S. stercoralis, так и S. ratti могут циклически проходить через одно свободноживущее поколение. Свободноживущее поколение состоит из постпаразитарных личинок, которые развиваются в свободноживущих взрослых самцов и самок; все потомство свободноживущих взрослых особей развивается в инфекционных личинок, которые должны заразить хозяина, чтобы продолжить жизненный цикл. Кроме того, этим экологическим или свободноживущим поколением можно экспериментально манипулировать в лаборатории. Поскольку свободноживущие взрослые strongyloides и взрослые особи C. elegans имеют схожую морфологию, такие методы, как интрагонадная микроинъекция, которые были первоначально разработаны для C. elegans, могут быть адаптированы для использования со свободноживущими взрослыми Strongyloides 4,5. В то время как ДНК обычно вводится в свободно живущих взрослых самок, как самцы, так и самки стронгилоидов могут быть микроинъективированы6. Таким образом, функциональные геномные инструменты доступны для изучения многих аспектов биологии стронгилоидов. Другие паразитические нематоды не имеют свободно живущего поколения и, как следствие, не так легко поддаются функциональным геномным методам3.
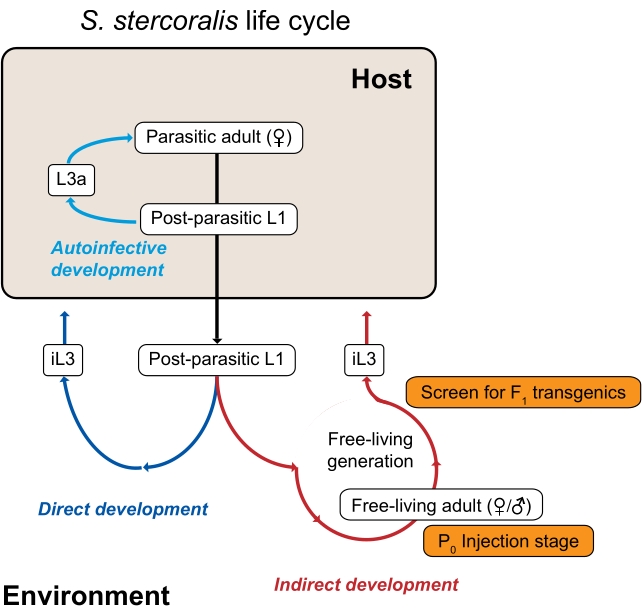
Рисунок 1: Жизненный цикл Strongyloides stercoralis. Паразитические самки S. stercoralis населяют тонкую кишку своих млекопитающих-хозяев (людей, нечеловеческих приматов, собак). Паразитические самки размножаются путем партеногенеза и откладывают яйца в тонком кишечнике. Яйца вылупляются, все еще находясь внутри хозяина, в постпаразитарные личинки, которые затем попадают в окружающую среду с фекалиями. Если постпаразитарные личинки являются самцами, они развиваются в свободноживущих взрослых самцов. Если постпаразитарные личинки являются самками, они могут либо развиться в свободноживущих взрослых самок (косвенное развитие), либо в личинок третьей стадии инфекции (iL3s; прямое развитие). Свободно живущие самцы и самки размножаются половым путем, чтобы создать потомство, которое ограничено, чтобы стать iL3s. При определенных условиях S. stercoralis также может подвергаться аутоинфекции, при которой часть постпаразитарных личинок остается внутри кишечника хозяина, а не попадает в окружающую среду с калом. Эти личинки могут развиваться в аутоинфективные личинки (L3a) внутри хозяина, проникать через стенку кишечника, мигрировать по организму и в конечном итоге возвращаться в кишечник, чтобы стать репродуктивными взрослыми. Жизненный цикл S. ratti аналогичен, за исключением того, что S. ratti заражает крыс и не имеет аутоинфекционного цикла. Генерация окружающей среды является ключом к использованию видов Strongyloides для генетических исследований. Свободно живущие взрослые самки (P0) могут быть микроинъективированы; их потомство, которое станет iL3s, является потенциальным трансгеномF1. Эта цифра была изменена по сравнению с Castelletto et al. 3. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
S. stercoralis разделяет многие аспекты своей биологии с другими желудочно-кишечными паразитическими нематодами, включая инвазию хозяина и иммунную модуляцию хозяина. Например, человеко-паразитические анкилостомы в родах Necator и Ancylostoma также заражаются при проникновении в кожу, аналогичным образом перемещаются по телу и в конечном итоге живут как паразитические взрослые в тонком кишечнике7. Таким образом, многие желудочно-кишечные нематоды, вероятно, используют общее сенсорное поведение и методы уклонения от иммунитета. В результате знания, полученные от Strongyloides , дополнят результаты у других менее генетически поддающихся лечению нематод и приведут к более полному пониманию этих сложных и важных паразитов.
Этот протокол микроинъекции описывает метод введения ДНК в Strongyloides свободноживущих взрослых самок для создания трансгенного и мутантного потомства. Описаны требования к поддержанию штамма, включая сроки развития взрослых червей для микроинъекций и сбор трансгенного потомства. Протоколы и демонстрация полной техники микроинъекции, наряду с протоколами культивирования и скрининга трансгенного потомства, включены вместе со списком всего необходимого оборудования и расходных материалов.
