Strongyloides stercoralis har længe været overset som et vigtigt humant patogen sammenlignet med de mere almindeligt anerkendte hageorm og rundorm Ascaris lumbricoides1. Tidligere undersøgelser af ormebyrden undervurderede ofte alvorligt forekomsten af S. stercoralis på grund af den lave følsomhed af almindelige diagnostiske metoder til S. stercoralis2. I de senere år har epidemiologiske undersøgelser baseret på forbedrede diagnostiske værktøjer anslået, at den sande forekomst af S. stercoralis-infektioner er meget højere end tidligere rapporteret, ca. 610 millioner mennesker over hele verden2.
Både S. stercoralis og andre Strongyloides-arter, herunder den nært beslægtede rotteparasit og fælles laboratoriemodel S. ratti, har en usædvanlig livscyklus, der er fordelagtig for eksperimentelle genomiske undersøgelser, fordi den består af både parasitære og fritlevende (miljømæssige) generationer3 (figur 1). Specifikt kan både S. stercoralis og S. ratti cykle gennem en enkelt fritlevende generation. Den fritlevende generation består af postparasitære larver, der udvikler sig til fritlevende voksne mænd og kvinder; alle afkom af de fritlevende voksne udvikler sig til infektiøse larver, som skal inficere en vært for at fortsætte livscyklussen. Desuden kan denne miljømæssige eller fritlevende generation eksperimentelt manipuleres i laboratoriet. Fordi fritlevende Strongyloides voksne og C. elegans voksne deler lignende morfologi, kan teknikker som intragonadal mikroinjektion, der oprindeligt blev udviklet til C. elegans, tilpasses til brug med fritlevende voksne Strongyloides 4,5. Mens DNA generelt introduceres i fritlevende voksne kvinder, kan både mænd og kvinder af Strongyloides mikroinjekt6. Således er funktionelle genomiske værktøjer tilgængelige for at forhøre mange aspekter af Strongyloidernes biologi. Andre parasitære nematoder mangler en fritlevende generation og er derfor ikke så let modtagelige for funktionelle genomiske teknikker3.
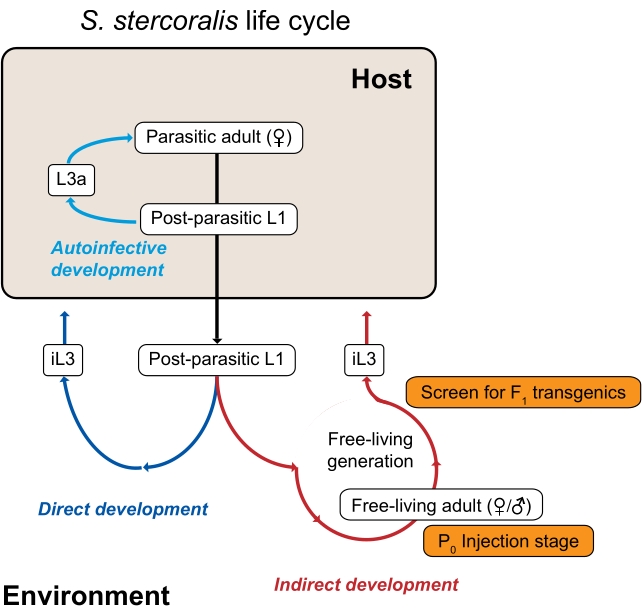
Figur 1: Strongyloides stercoralis livscyklus. S. stercoralis parasitære kvinder beboer tyndtarmen hos deres pattedyrværter (mennesker, ikke-menneskelige primater, hunde). De parasitære hunner reproducerer ved parthenogenese og lægger æg i tyndtarmen. Æggene klækkes, mens de stadig er inde i værten, til postparasitære larver, som derefter overføres til miljøet med afføring. Hvis de postparasitære larver er mandlige, udvikler de sig til fritlevende voksne mænd. Hvis de postparasitære larver er kvindelige, kan de enten udvikle sig til fritlevende voksne hunner (indirekte udvikling) eller tredje fase infektiøse larver (iL3s; direkte udvikling). De fritlevende mænd og kvinder reproducerer seksuelt for at skabe afkom, der er begrænset til at blive iL3’er. Under visse betingelser kan S. stercoralis også gennemgå autoinfektion, hvor nogle af de postparasitære larver forbliver inde i værtstarmen i stedet for at passere ind i miljøet i afføring. Disse larver kan udvikle sig til autoinfektive larver (L3a) inde i værten, trænge gennem tarmvæggen, migrere gennem kroppen og til sidst vende tilbage til tarmen for at blive reproduktive voksne. Livscyklussen for S. ratti er ens, bortset fra at S. ratti inficerer rotter og ikke har en autoinfektiv cyklus. Miljøgenerationen er nøglen til at bruge Strongyloides-arter til genetiske undersøgelser. De fritlevende voksne kvinder (P0) kan mikroinjekteres; deres afkom, som alle bliver iL3’er, er de potentielle F1-transgene. Dette tal er blevet ændret fra Castelletto et al. 3. Klik her for at se en større version af denne figur.
S. stercoralis deler mange aspekter af sin biologi med andre gastrointestinale human-parasitiske nematoder, herunder værtsinvasion og værtsimmunmodulation. For eksempel inficerer human-parasitiske hookworms i slægterne Necator og Ancylostoma også ved hudindtrængning, navigerer på samme måde gennem kroppen og i sidste ende befinder sig som parasitære voksne i tyndtarmen7. Således bruger mange gastrointestinale nematoder sandsynligvis almindelig sensorisk adfærd og immununddragelsesteknikker. Som følge heraf vil den viden, der er hentet fra Strongyloides , supplere fund i andre mindre genetisk medgørlige nematoder og føre til en mere fuldstændig forståelse af disse komplekse og vigtige parasitter.
Denne mikroinjektionsprotokol skitserer metoden til at introducere DNA i Strongyloides fritlevende voksne kvinder for at lave transgene og mutante afkom. Kravene til vedligeholdelse af stammen, herunder udviklingstimingen af voksne orme til mikroinjektioner og indsamling af transgene afkom, er beskrevet. Protokoller og en demonstration af den komplette mikroinjektionsteknik sammen med protokoller til dyrkning og screening af transgene afkom er inkluderet sammen med en liste over alt nødvendigt udstyr og forbrugsstoffer.
