Образование кости остеобластом сопровождается скоординированным разрушением или рассасыванием костей остеокластами. Баланс между остеобластным формированием кости и резорбцией остеокластов представляет собой связанный процесс, описывающий оборот или ремоделирование кости, который необходим для скелетного гомеостаза. Дисфункция остеобластов приводит к нарушению костеобразования и приводит к различным заболеваниям, в том числеостеопорозу 1,2,3. Дифференцировка стромальных стволовых клеток костного мозга (BMSCs) ex vivo/in vitro к предшественникам остеобластов и зрелым остеобластам приводит к образованию и отложению минерализованного костного матрикса в культуральном сосуде с течением времени 4,5,6. Это костное образование остеобластом требует значительного количества клеточной энергии. В частности, было показано, что синтез и секреция коллагена в значительной степени зависят от клеточных соотношений АТФ: АДФ, и, по-видимому, минерализованный пузырчатый трафик и секреция требуют дополнительного АТФ 7,8,9,10,11. Многие исследователи продемонстрировали, что процесс остеобластогенеза и функции остеобластов требует адекватного запаса энергии для удовлетворения метаболической потребности в формировании костей 12,13,14,15,16. Поэтому целью данного метода является характеристика биоэнергетического статуса первичных, мышиных стромальных клеток на протяжении дифференцировки остеобластов с помощью анализатора метаболического потока клеток реального времени. Эти методы помогают в развитии лучшего понимания скелетного гомеостаза, что в конечном итоге может привести к разработке новых терапевтических вариантов, способных улучшить скелетные расстройства.
Анализатор метаболического потока клеток в реальном времени может быть использован для измерения скорости потребления кислорода (OCR) и скорости внеклеточного подкисления (ECAR) живых остеобластов, что соответствует митохондриальной и гликолитической продукции АТФ соответственно. Основополагающим для этой методологии является тот факт, что один ион H+ на лактат высвобождается во время гликолиза при превращении глюкозы в лактат, что изменяет рН среды, отраженный в значениях ECAR. И наоборот, во время цикла TCA (трикарбоновой кислоты) окислительное фосфорилирование через митохондрии производит CO2 путем использования или потребления кислорода, и поэтому мониторинг OCR отражает этот метаболический процесс. Анализатор измеряет как OCR, так и ECAR во внеклеточной микросреде одновременно и в режиме реального времени, что позволяет использовать огромный потенциал при изучении клеточной биоэнергетики 6,17. Кроме того, выполнение этих анализов относительно просто и легко настраивается в зависимости от экспериментальной цели. Аналогичные методы были использованы для дальнейшего понимания метаболической регуляции Т-клеток иммунной системы18,19, инициации рака и прогрессирования20, наряду с несколькими другими типами клеток, способствующими метаболическим синдромам21,22.
Преимущества анализатора метаболических потоков в реальном времени перед альтернативными методами включают (1) возможность измерения клеточной биоэнергетики живых клеток в режиме реального времени, (2) способность выполнять анализ с относительно небольшим количеством клеток (требуется всего 5000 клеток), (3) инъекционные порты для параллельного манипулирования несколькими обработками в высокопроизводительной 96-луночной системе, (4) использование автоматического тепловизора без радиоактивных меток для нормализации18, 23,24. Следующие методы направлены на обобщенное, но подробное описание мониторинга клеточной биоэнергетики в мышиных BMSC на протяжении дифференцировки остеобластов с помощью анализатора. Он будет включать в себя регулярно выполняемые анализы; однако, как и во многих техниках и методах, настоятельно рекомендуется, чтобы отдельные лаборатории определяли конкретные детали для своих экспериментов.
Выбор анализов и различные типы анализов доступны: Доступно большое разнообразие наборов для анализа и реагентов для изучения биоэнергетики клеток, обеспечивая при этом надежность и согласованность экспериментальных результатов. Кроме того, настольное программное обеспечение также предлагает шаблоны анализа, которые можно легко настроить. Анализ может быть определен на основе потребностей пользователя в измерении различных метаболических параметров. Эти анализы могут быть модифицированы различными способами на основе экспериментальной цели и/или научного вопроса. Например, с четырьмя инъекционными портами несколько соединений могут быть введены в пробирную среду для анализа клеточного ответа, специфичного для каждого метаболического пути.
Тест фенотипа клеточной энергии: Этот анализ измеряет метаболический фенотип живых клеток и метаболический потенциал. Этот анализ также рекомендуется в качестве первого шага для получения обобщенного представления о метаболизме, специфичном для пути. Смесь олигомицина А-ингибитора АТФ-синтазы и карбонилцианида 4-(трифторметокси)фенилгидразона (FCCP)-митохондриального разъединяющего агента вводят для понимания энергетического потенциала клетки. Инъекция олигомицина А ингибирует синтез АТФ, что приводит к увеличению скорости гликолиза (ЭКАР), чтобы позволить клеткам удовлетворить свои энергетические потребности; с другой стороны, инъекция FCCP приводит к более высокому OCR из-за деполяризации митохондриальной мембраны. По сути, этот анализ изображает базальное метаболическое дыхание и после двойных инъекций, толчков или стрессов метаболическую реакцию. Основываясь на этих параметрах, программное обеспечение затем строит ГРАФИК OCR и ECAR клеток, классифицируя клетки как аэробное, покоящееся, гликолитическое или энергетическое состояние с течением времени25,26.
Анализ скорости производства АТФ в режиме реального времени: Это измеряет выработку клеточной АТФ одновременно от гликолиза и митохондриального дыхания. Этот анализ количественно измеряет метаболические сдвиги от двух энергетических путей и предоставляет данные о скорости производства митохондриальной и гликолитической АТФ с течением времени. Анализ получает базальные данные OCR и ECAR с последующим расчетом скорости производства митохондриальной АТФ путем инъекции олигомицина А и гликолитической скорости производства АТФ путем инъекции смеси ротенона + антимицина А (полное ингибирование митохондриальной функции), что приводит к митохондриальному подкислению17,27.
Клеточный митохондриальный стресс-тест (или клеточный мито-стресс-тест): Он измеряет митохондриальную функцию через АТФ-связанное дыхание, количественно оценивает клеточную биоэнергетику, идентифицирует митохондриальную дисфункцию и измеряет реакцию клеток на стресс. Различные параметры, включая базальную и запасную дыхательную способность, АТФ-связанное дыхание, максимальное дыхание и немитохондриальное потребление кислорода, могут быть получены в одном анализе. Этот анализ включает последовательные инъекции олигомицина А, FCCP (митохондриального разъединяющего агента), смеси ингибиторов ротенона/ антимицина А для эффективного анализа их влияния на митохондриальную функцию28.
Тест на гибкость топлива: Это измеряет скорость митохондриального дыхания путем окисления трех основных митохондриальных топлив присутствием и отсутствием их ингибиторов. Последовательное ингибирование глюкозы, глутамина и жирных кислот помогает измерить зависимость, емкость и гибкость клеток, а также зависимость клеток в различных клеточных путях для удовлетворения потребности в энергии. Когда митохондрии не могут удовлетворить требования заблокированного пути, представляющего интерес, путем окисления других видов топлива, клетки входят в состояние зависимости. Емкость клеток рассчитывается путем ингибирования двух других альтернативных путей с последующим ингибированием интересующего пути. Гибкость клеток помогает понять способность митохондрий компенсировать и удовлетворять потребности в топливе ингибированного пути. Он рассчитывается путем вычитания зависимости ячеек из емкости ячеек. Три различных ингибитора используются независимо или в виде смеси двух для эффективного расчета параметров анализа. 2-циан-3-(1-фенил-1Н-индол-3-ил)-2-пропеновая кислота (UK5099) ингибирует окисление глюкозы, блокируя пируват-носитель в гликолизе. Бис-2-(5-фенилацетамидо-1,3,4-тиадиазол-2-ил) (BPTES) этилсульфид ингибирует путь окисления глутамина, а этомоксир ингибирует окисление длинноцепочечных жирных кислот29.
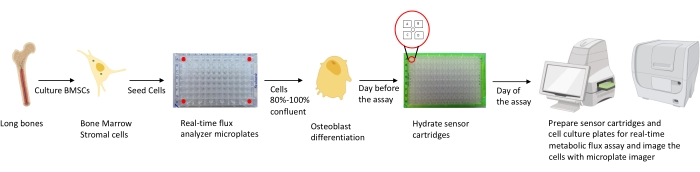
Рисунок 1: Схематическое представление методологии культивирования и подготовки остеобластов к анализу. Мышиные BMSC выделяют из длинных костей, культивируют и засеивают в 96-луночные пластины с плотностью 25 000 клеток на лунку. Культивирование этих клеток в специфических средах остеобластов начинается, когда они достигают 80%-100% конфлюзии, чтобы начать их дифференцировку. Анализы выполняются на разных этапах дифференцировки. Картриджные пластины гидратируются за один день до анализа. В день анализа различные ингибиторы вводятся в порты картриджей датчиков в зависимости от требований анализа, а к калибровочной пластине на 96 скважин добавляется калибровочный буфер. После калибровки выполняется анализ метаболического потока клеток в реальном времени, после чего выполняется визуализация микропластины клеточной культуры с использованием микропластичного тепловизора для нормализации данных анализатора метаболического потока клеток в реальном времени с количеством клеток. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
