La plupart des protéines eucaryotes complexes subissent des modifications post-traductionnelles élaborées après leur expression, nécessitant un repliement des protéines hautement assisté et des cofacteurs pour être fonctionnels1. La production de grandes quantités de protéines humaines solubles dans un hôte bactérien reste un défi important en raison des coûts élevés et du manque de méthodes d’expression et de purification robustes, même pour des expériences de laboratoire à plus petite échelle2,3. Les MMP, des endopeptidases humaines de grand poids moléculaire, sont généralement exprimées sous forme de corps d’inclusion insolubles lorsqu’elles sont exprimées dans E. coli. L’extraction des MMP humains solubles conduit souvent à un processus de solubilisation et de repli laborieux et long4.
Les MMP jouent un rôle essentiel dans les processus physiologiques et pathogènes. Les MMP humains sont une famille de 23 endopeptidases de zinc, classées par structure et spécificité du substrat, et exprimées différemment en dépit d’un domaine catalytique hautement conservé5,6. Les MMP sont sécrétés sous forme de zymogènes inactifs, régulés par activation post-traductionnelle et leurs inhibiteurs endogènes, inhibiteurs tissulaires des métalloprotéinases (TIMPs)7,8,9,10. Bien qu’initialement reconnus pour leur rôle dans le renouvellement de l’ECM, les MMP ont également été impliqués dans le développement, la morphogenèse, la réparation des tissus et le remodelage8. La dérégulation des MMP a été notamment liée au cancer ainsi qu’aux maladies neurodégénératives, cardiovasculaires et fibrotiques, entre autres maladies5,7.
Le développement de méthodes robustes de production de MMP à grande échelle est essentiel pour assurer le succès des études futures des mécanismes MMP par le biais de tests biochimiques et cellulaires. Divers MMP ont déjà été exprimés dans des bactéries11, y compris des MMP marqués Hisx6, sans modifier l’activité MMP12,13,14,15. Cependant, ces méthodes comprennent des étapes longues et fastidieuses qui peuvent être difficiles à reproduire.
Les cellules de mammifères peuvent également être utilisées pour exprimer de nombreuses protéines humaines différentes tout en assurant les modifications post-traductionnelles appropriées16. Bien que le système d’expression des mammifères soit un choix idéal pour produire des protéines humaines recombinantes avec des modifications post-traductionnelles appropriées, les principaux inconvénients de cette méthode sont les faibles rendements initiaux, les milieux de croissance et les réactifs coûteux, les longs délais pour atteindre des lignes d’expression stables et le risque de contamination par d’autres espèces telles que les champignons ou les bactéries2,11 . De plus, la production de MMP dans les lignées cellulaires de mammifères produit des impuretés à partir de protéines cellulaires associées telles que les TIMPs ou les fibronectines11. Contrairement à la croissance cellulaire lente observée dans les cellules de mammifères, le système d’expression bactérienne offre une production de protéines à grande échelle dans un court laps de temps ainsi que des milieux plus simples et des besoins de croissance. Cependant, en raison de l’absence d’autres protéines cellulaires associées (c.-à-d. les TIMPs) dans les systèmes d’expression bactérienne, les MMP actifs à des concentrations plus élevées sont sujets à la dégradation par autoprotéolyse, ce qui entraîne un faible rendement en MMP17.
Cet article décrit une méthode détaillée pour l’expression bactérienne, la purification et l’activation de Hisx6-pro-MMP-3cd recombinant en utilisant E. coli comme hôte d’expression en raison de son prix abordable, de sa simplicité et de son succès dans la production de rendements plus élevés de MMP2,3,18. Étant donné qu’E. coli n’a pas la machinerie de repliement des protéines et le traitement post-traductionnel requis pour les MMP recombinants et d’autres protéines complexes, de nombreuses souches d’E. coli ont été conçues pour surmonter ces limitations, faisant d’E. coli un hôte plus approprié pour l’expression de MMP-3cd humain recombinant,19,20 . Par exemple, la souche R2DP utilisée dans cette étude améliore l’expression eucaryote en fournissant un plasmide résistant au chloramphénicol contenant des codons rarement utilisés chez E. coli.
Comme décrit dans ce protocole, après surexpression de corps d’inclusion relativement purs du vecteur pET-3a (Figure 1) dans les cellules R2DP, les protéines du domaine catalytique Hisx6-pro-MMP-3 (MMP-3cd) sont extraites et dénaturées4. Hisx6-pro-MMP-3cd3,19 a été purifié par chromatographie par étiquette d’affinité. Lors du repliement et de la dialyse, le pro-MMP-3cd (zymogène) a été activé par l’acétate de 4-aminophénylmercurique (APMA), et l’analyse SDS-PAGE est utilisée pour évaluer les rendements et la nécessité d’une purification supplémentaire5,21. Ce protocole décrit l’expression, la purification et l’activation de MMP-3cd soluble à titre d’exemple. Cependant, il peut également être utilisé comme guide pour l’expression d’autres MMP et protéases humaines ayant une expression similaire, et des mécanismes d’activation (Figure 2). Pour d’autres protéines autres que MMP-3cd, il est conseillé au lecteur de déterminer les compositions tampons optimales et les méthodes pour leur protéine cible avant d’essayer ce protocole.
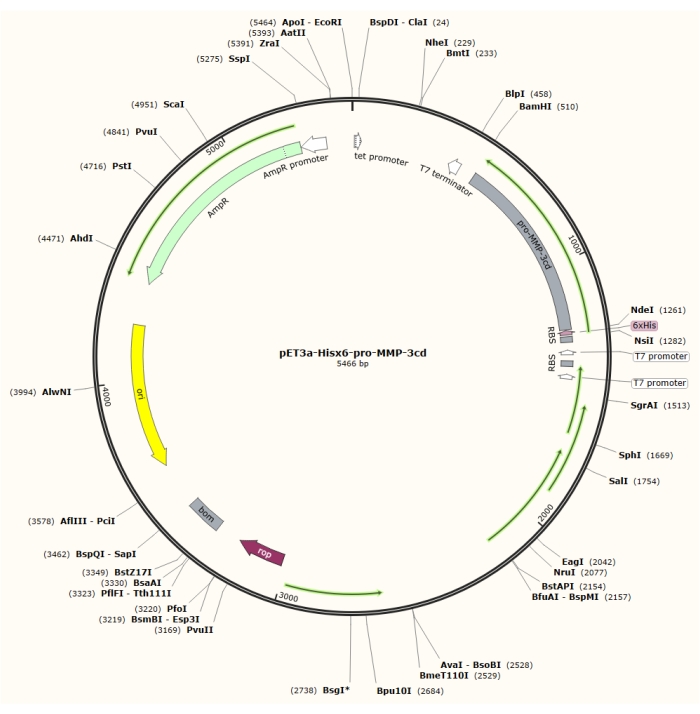
Figure 1 : Carte plasmidique du plasmide pET-3a-Hisx6-pro-MMP-3cd. Le vecteur pET-3a comprend un gène de résistance à l’ampicilline. Une séquence de balises Hisx6 n-terminales est clonée dans le vecteur pET-3a, y compris pro-MMP-3cd, pour donner la construction pET-3a-Hisx6-pro-MMP-3cd sous contrôle du promoteur T7 entre les sites de restriction BamHI et NdeI. Veuillez cliquer ici pour voir une version agrandie de cette figure.
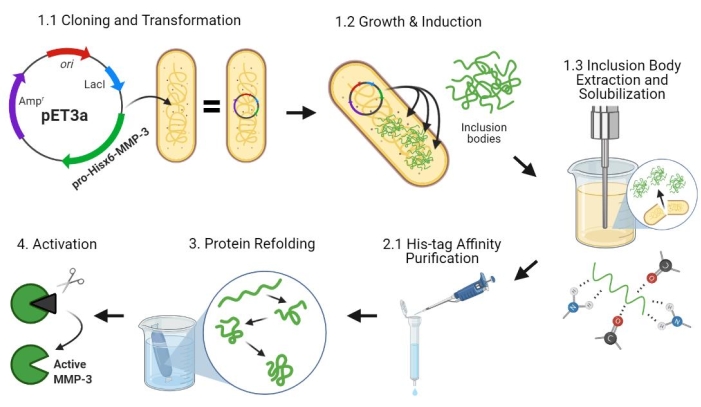
Figure 2 : Expression bactérienne du pro-MMP-3cd, purification, repliement et activation. 1.1 : Le plasmide pET-3a-Hisx6-pro-MMP-3cd a été transformé en cellules BL21(DE3) ou R2DP. 1.2 : L’expression de la protéine Pro-MMP-3cd a été induite à l’aide de l’IPTG. 1.3: La lyse chimique et la sonication sont utilisées pour extraire les protéines Hisx6-pro-MMP-3cd qui sont principalement insolubles et présentes dans les corps d’inclusion. L’urée a été utilisée pour dénaturer et solubiliser les protéines des corps d’inclusion. 2.1. La protéine Hisx6-pro-MMP-3cd dénaturée a été purifiée par purification par chromatographie d’affinité. 3. L’Hisx6-pro-MMP-3cd élué a été lentement replié pendant la dialyse par élimination progressive de l’urée du tampon. 4. Enfin, la protéine MMP-3cd repliée a été activée à l’aide de l’APMA en supprimant le domaine pro-peptidique N-terminal. L’APMA est ensuite retiré de la solution par dessalement. Les numéros correspondent aux sections du protocole décrivant ces étapes. Abréviations : MMP-3cd = Matrice métalloprotéinase-3 domaine catalytique ; APMA = acétate de 4-aminophénylmercurique. Veuillez cliquer ici pour voir une version agrandie de cette figure.
