Die meisten komplexen eukaryotischen Proteine durchlaufen nach der Expression ausgeklügelte posttranslationale Modifikationen, die eine hochunterstützte Proteinfaltung und Co-Faktoren erfordern, um funktionsfähig zu sein1. Die Herstellung großer Mengen löslichen menschlichen Proteins in einem bakteriellen Wirt bleibt aufgrund der hohen Kosten und des Mangels an robusten Expressions- und Reinigungsmethoden eine erhebliche Herausforderung, selbst für kleinere Laborexperimente2,3. MMPs, humane Endopeptidasen mit großem Molekulargewicht, werden normalerweise als unlösliche Einschlusskörper exprimiert, wenn sie in E. coli exprimiert werden. Die Extraktion von löslichen humanen MMPs führt oft zu einem mühsamen, zeitaufwändigen Solubilisierungs- und Umfaltungsprozess4.
MMPs spielen eine entscheidende Rolle sowohl in physiologischen als auch in pathogenen Prozessen. Humane MMPs sind eine Familie von 23 Zink-Endopeptidasen, kategorisiert nach Struktur- und Substratspezifität und differentiell exprimiert trotz einer hochkonservierten katalytischen Domäne5,6. MMPs werden als inaktive Zymogene sezerniert, reguliert durch posttranslationale Aktivierung und ihre endogenen Inhibitoren, Gewebeinhibitoren von Metalloproteinasen (TIMPs)7,8,9,10. Obwohl MMPs ursprünglich für ihre Rolle beim ECM-Umsatz anerkannt waren, waren sie auch an der Entwicklung, Morphogenese, Gewebereparatur und -umgestaltung beteiligt8. Die Dysregulation von MMPs wurde unter anderem mit Krebs sowie neurodegenerativen, kardiovaskulären und fibrotischen Erkrankungen in Verbindung gebracht5,7.
Die Entwicklung robuster MMP-Produktionsmethoden im großen Maßstab ist entscheidend, um den Erfolg zukünftiger Studien von MMP-Mechanismen durch biochemische und zellbasierte Assays sicherzustellen. Verschiedene MMPs wurden zuvor in Bakterien exprimiert11, einschließlich Hisx6-markierter MMPs, ohne die MMP-Aktivität zu verändern12,13,14,15. Diese Methoden umfassen jedoch mühsame, lange Schritte, die möglicherweise schwer zu replizieren sind.
Säugetierzellen können auch verwendet werden, um viele verschiedene menschliche Proteine zu exprimieren und gleichzeitig die richtigen posttranslationalen Modifikationen zu gewährleisten16. Obwohl das Expressionssystem für Säugetiere eine ideale Wahl ist, um rekombinante menschliche Proteine mit geeigneten posttranslationalen Modifikationen herzustellen, sind die Hauptnachteile dieser Methode anfänglich niedrige Ausbeuten, kostspielige Wachstumsmedien und Reagenzien, lange Zeiträume zum Erreichen stabiler Mimiklinien und das Risiko einer Kontamination mit anderen Spezies wie Pilzen oder Bakterien2,11 . Darüber hinaus führt die MMP-Produktion in Zelllinien von Säugetieren zu Verunreinigungen aus assoziierten zellulären Proteinen wie TIMPs oder Fibronektinen11. Im Gegensatz zu dem langsamen Zellwachstum, das in Säugetierzellen beobachtet wird, bietet das bakterielle Expressionssystem eine groß angelegte Proteinproduktion in kurzer Zeit sowie einfachere Medien- und Wachstumsanforderungen. Aufgrund des Fehlens anderer assoziierter zellulärer Proteine (d. h. TIMPs) in bakteriellen Expressionssystemen unterliegen aktive MMPs in höheren Konzentrationen jedoch dem Abbau durch Autoproteolyse, was zu einer schlechten MMP-Ausbeute führt17.
Dieses Papier beschreibt eine detaillierte Methode zur bakteriellen Expression, Reinigung und Aktivierung von rekombinantem Hisx6-pro-MMP-3cd unter Verwendung von E. coli als Ausdruckshost aufgrund seiner Erschwinglichkeit, Einfachheit und seines Erfolgs bei der Erzeugung höherer Ausbeuten von MMPs2,3,18. Da E. coli nicht über die Proteinfaltungsmaschinerie und die posttranslationale Verarbeitung verfügt, die für rekombinante MMPs und andere komplexe Proteine erforderlich sind, wurden viele E. coli-Stämme entwickelt, um diese Einschränkungen zu überwinden, was E. coli zu einem geeigneteren Wirt für die Expression von rekombinantem humanem MMP-3cd macht,19,20 . Zum Beispiel verbessert der in dieser Studie verwendete R2DP-Stamm die eukaryotische Expression, indem er ein chloramphenicolresistentes Plasmid liefert, das Codons enthält, die selten in E. coli verwendet werden.
Wie in diesem Protokoll beschrieben, werden nach Überexpression von relativ reinen Einschlusskörpern aus dem pET-3a-Vektor (Abbildung 1) in R2DP-Zellen Hisx6-pro-MMP-3-Proteine (MMP-3cd) extrahiert und denaturiert4. Hisx6-pro-MMP-3cd3,19 wurde mittels Affinitäts-Tag-Chromatographie gereinigt. Nach der Rückfaltung und Dialyse wurde das Pro-MMP-3cd (Zymogen) durch 4-Aminophenylquecksilberacetat (APMA) aktiviert, und die SDS-PAGE-Analyse wird verwendet, um die Ausbeuten und die Notwendigkeit einer weiteren Reinigung zu bewerten5,21. Dieses Protokoll beschreibt als Beispiel den Ausdruck, die Reinigung und die Aktivierung von löslichem MMP-3cd. Es kann jedoch auch als Leitfaden für die Expression anderer MMPs und menschlicher Proteasen mit ähnlicher Expression und Aktivierungsmechanismen verwendet werden (Abbildung 2). Für andere Proteine als MMP-3cd wird dem Leser empfohlen, optimale Pufferzusammensetzungen und -methoden für sein Zielprotein zu bestimmen, bevor er dieses Protokoll versucht.
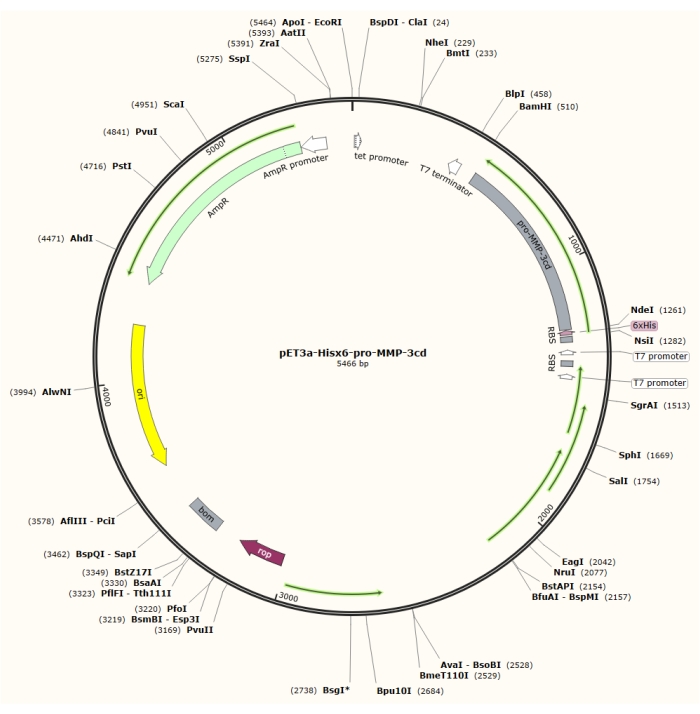
Abbildung 1: Plasmidabbildung des pET-3a-Hisx6-pro-MMP-3cd-Plasmids. Der pET-3a-Vektor enthält ein Ampicillin-Resistenz-Gen. Eine N-terminale Hisx6-Tag-Sequenz wird in den pET-3a-basierten Vektor, einschließlich pro-MMP-3cd, geklont, um das pET-3a-Hisx6-pro-MMP-3cd-Konstrukt unter Kontrolle des T7-Promotors zwischen BamHI- und NdeI-Restriktionssites zu erhalten. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
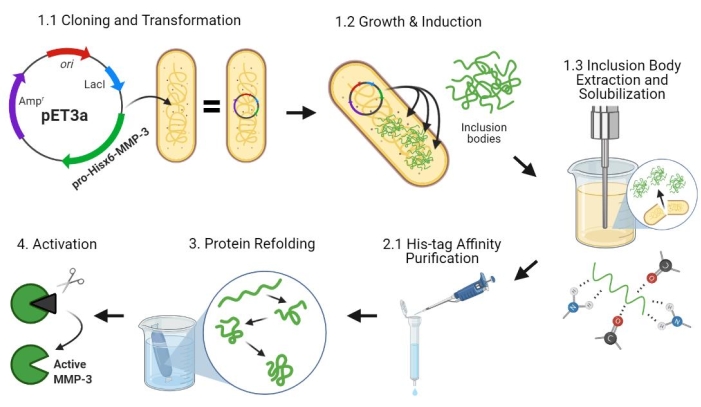
Abbildung 2: Bakterielle Expression von Pro-MMP-3cd, Reinigung, Rückfaltung und Aktivierung . 1.1: pET-3a-Hisx6-pro-MMP-3cd-Plasmid wurde in BL21(DE3)- oder R2DP-Zellen umgewandelt. 1.2: Die Pro-MMP-3cd-Proteinexpression wurde mittels IPTG induziert. 1.3: Chemische Lyse und Beschallung werden verwendet, um Hisx6-pro-MMP-3cd-Proteine zu extrahieren, die hauptsächlich unlöslich sind und in den Einschlusskörpern vorkommen. Harnstoff wurde verwendet, um Protein aus Einschlusskörpern zu denaturieren und zu lösen. 2.1. Das denaturierte Hisx6-pro-MMP-3cd-Protein wurde durch affinitätschromatographische Reinigung gereinigt. 3. Die eluierte Hisx6-pro-MMP-3cd wurde während der Dialyse durch allmähliche Entfernung von Harnstoff aus dem Puffer langsam wieder gefaltet. 4. Schließlich wurde das gefaltete MMP-3cd-Protein mit APMA aktiviert, indem die N-terminale Pro-Peptid-Domäne entfernt wurde. APMA wird später durch Entsalzen aus der Lösung entfernt. Die Zahlen entsprechen den Protokollabschnitten, die diese Schritte beschreiben. Abkürzungen: MMP-3cd = Matrix-Metalloproteinase-3-katalytische Domäne; APMA = 4-Aminophenylquecksilberacetat. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
