De fleste komplekse eukaryote proteiner gjennomgår forseggjorte posttranslasjonelle modifikasjoner etter uttrykk, noe som krever svært assistert proteinfolding og kofaktorer for å være funksjonelle1. Å produsere store mengder løselig humant protein i en bakteriell vert er fortsatt en betydelig utfordring på grunn av høye kostnader og mangel på robuste uttrykks- og rensemetoder, selv for mindre laboratorieforsøk2,3. MLP-er, humane endopeptidaser med stor molekylvekt, uttrykkes vanligvis som uoppløselige inklusjonslegemer når de uttrykkes i E. coli. Utvinning av løselige menneskelige MMP-er fører ofte til en arbeidskrevende, tidkrevende løsnings- og omfoldingsprosess4.
MMP-er har kritiske roller i både fysiologiske og patogene prosesser. Humane MMP-er er en familie på 23 sink endopeptidaser, kategorisert etter struktur og substratspesifisitet, og differensialt uttrykt til tross for et høyt bevart katalytisk domene5,6. MMP-er utskilles som inaktive zymogener, regulert via posttranslasjonell aktivering og deres endogene hemmere, vevshemmere av metalloproteinaser (TIMPs) 7,8,9,10. Selv om de først ble anerkjent for sin rolle i ECM-omsetning, har MMP-er også vært involvert i utvikling, morfogenese, vevsreparasjon og ombygging8. Dysregulering av MMP-er har særlig vært knyttet til kreft sammen med blant annet nevrodegenerative, kardiovaskulære og fibrotiske sykdommer.
Utviklingen av robuste MMP-produksjonsmetoder i stor skala er avgjørende for å sikre suksess for fremtidige studier av MMP-mekanismer gjennom biokjemiske og cellebaserte analyser. Ulike MMP-er har tidligere blitt uttrykt i bakterier11, inkludert Hisx6-taggede MMPer, uten å endre MMP-aktivitet12,13,14,15. Imidlertid inkluderer disse metodene kjedelige, lange trinn som kan være vanskelige å gjenskape.
Pattedyrceller kan også brukes til å uttrykke mange forskjellige menneskelige proteiner samtidig som de sikrer riktige posttranslasjonelle modifikasjoner16. Selv om pattedyruttrykkssystemet er et ideelt valg for å produsere rekombinante menneskelige proteiner med riktige postoversettelsesendringer, er de viktigste ulempene ved denne metoden innledende lave utbytter, kostbare vekstmedier og reagenser, lange tidslinjer for å nå stabile uttrykkslinjer og risiko for forurensning med andre arter som sopp eller bakterier2,11 . Videre gir MMP-produksjon i pattedyrcellelinjer urenheter fra tilknyttede cellulære proteiner som TIMPer eller fibronectins11. I motsetning til den langsomme celleveksten som observeres i pattedyrceller, tilbyr bakterieuttrykkssystemet storskala proteinproduksjon på kort tid sammen med enklere medier og vekstkrav. På grunn av mangel på andre assosierte cellulære proteiner (dvs. TIMPer) i bakterielle uttrykkssystemer, er aktive MMP-er ved høyere konsentrasjoner utsatt for nedbrytning gjennom autoproteolyse, noe som resulterer i dårlig MMP-utbytte17.
Dette dokumentet beskriver en detaljert metode for bakteriell uttrykk, rensing og aktivering av rekombinant Hisx6-pro-MMP-3cd ved hjelp av E. coli som uttrykksvert på grunn av sin overkommelige pris, enkelhet og suksess med å produsere høyere utbytter av MMPs2,3,18. Siden E. coli mangler protein folding maskiner og posttranslational behandling som kreves for rekombinante MMP og andre komplekse proteiner, mange E. coli stammer har blitt konstruert for å overvinne disse begrensningene, noe som gjør E. coli en mer egnet vert for uttrykk for rekombinant menneskelig MMP-3cd, 19,20 . For eksempel forbedrer R2DP-stammen som brukes i denne studien eukaryotisk uttrykk ved å levere en kloramfenikolresistent plasmid som inneholder codons som sjelden brukes i E. coli.
Som beskrevet i denne protokollen, etter overekspressering av relativt rene inklusjonslegemer fra pET-3a-vektoren (figur 1) i R2DP-celler, blir Hisx6-pro-MMP-3 katalytiske domene (MMP-3cd) proteiner ekstrahert og denaturert4. Hisx6-pro-MMP-3cd3,19 ble renset ved hjelp av affinitetsmerkekromatografi. Ved omfolding og dialyse ble pro-MMP-3cd (zymogen) aktivert av 4-aminofenylmercuric acetat (APMA), og SDS-PAGE analyse brukes til å evaluere utbytter og behovet for ytterligere rensing5,21. Denne protokollen beskriver uttrykk, rensing og aktivering av løselig MMP-3cd som et eksempel. Det kan imidlertid også brukes som en veiledning for uttrykk for andre MMP-er og menneskelige proteaser med lignende uttrykk, og aktiveringsmekanismer (figur 2). For andre proteiner enn MMP-3cd anbefales leseren å bestemme optimale buffersammensetninger og metoder for deres målprotein før du prøver denne protokollen.
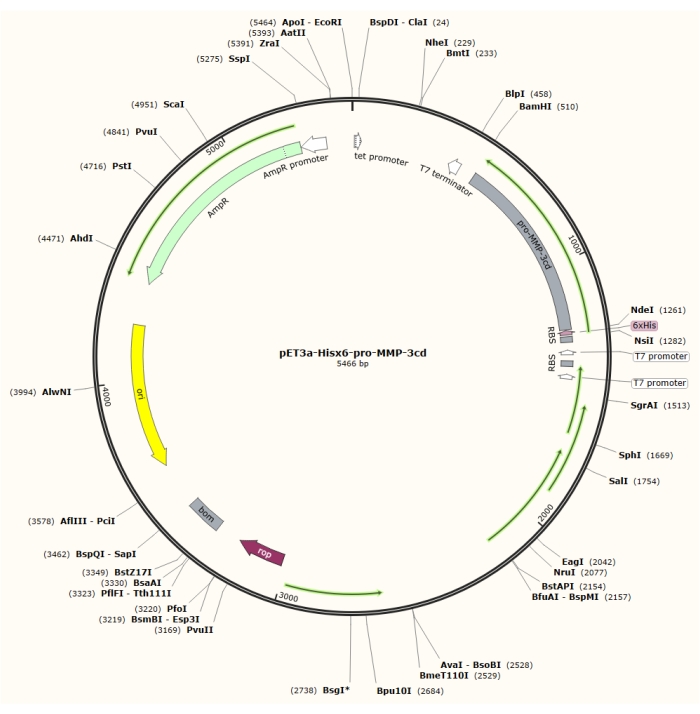
Figur 1: Plasmidkart over pET-3a-Hisx6-pro-MMP-3cd plasmid. PET-3a-vektoren inneholder et ampicillinresistensgen. En N-terminal Hisx6-tag-sekvens klones inn i den pET-3a-baserte vektoren, inkludert pro-MMP-3cd, for å gi pET-3a-Hisx6-pro-MMP-3cd-konstruksjonen under kontroll av T7-promotoren mellom BamHI og NdeI-begrensningssteder. Klikk her for å se en større versjon av denne figuren.
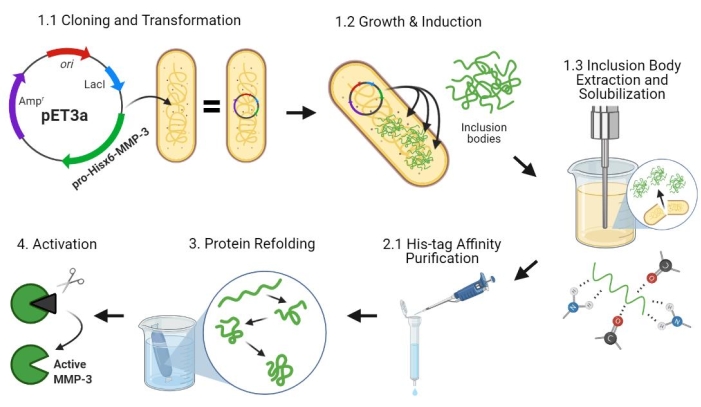
Figur 2: Bakteriell uttrykk for pro-MMP-3cd, rensing, bretting og aktivering. 1.1: pET-3a-Hisx6-pro-MMP-3cd plasmid ble forvandlet til BL21(DE3) eller R2DP Cells. 1.2: Pro-MMP-3cd proteinuttrykk ble indusert ved hjelp av IPTG. 1.3: Kjemisk lysis og sonikering brukes til å trekke ut Hisx6-pro-MMP-3cd proteiner som hovedsakelig er uoppløselige og finnes i inklusjonslegemene. Urea ble brukt til å denaturere og løse protein fra inklusjonslegemer. 2.1. Denaturert Hisx6-pro-MMP-3cd protein ble renset via affinitet kromatografi rensing. 3. Den eluted Hisx6-pro-MMP-3cd ble sakte omdelt under dialyse gjennom gradvis fjerning av urea fra bufferen. 4. Til slutt ble refoldet MMP-3cd protein aktivert ved hjelp av APMA ved å fjerne N-terminal pro-peptiddomenet. APMA fjernes senere fra løsningen gjennom avsalting. Tallene tilsvarer protokolldelene som beskriver disse trinnene. Forkortelser: MMP-3cd = Matrix metalloproteinase-3 katalytisk domene; APMA = 4-aminofenylmercuric acetat. Klikk her for å se en større versjon av denne figuren.
