Die Linse ist ein klares und eiförmiges Gewebe in der vorderen Augenkammer, das aus zwei Zelltypen besteht, Epithel- und Faserzellen 1 (Abbildung 1). Es gibt eine Monoschicht von Epithelzellen, die die vordere Hemisphäre der Linse bedeckt. Faserzellen werden von Epithelzellen unterschieden und machen den Großteil der Linse aus. Die hochspezialisierten Faserzellen durchlaufen eine Elongations-, Differenzierungs- und Reifungsprogrammierung, die durch deutliche Veränderungen der Zellmembranmorphologie von der Linsenperipherie bis zum Linsenzentrum gekennzeichnetist 2,3,4,5,6,7,8,9,10,11,12 , auch bekannt als Linsenkern. Die Funktion der Linse, Licht aus verschiedenen Entfernungen auf die Netzhaut zu fokussieren, hängt von ihren biomechanischen Eigenschaften ab, einschließlich Steifigkeit und Elastizität 13,14,15,16,17,18,19. Die komplexen Verzahnungen von Linsenfasern wurden hypothetisch aufgestellt20,21 und haben kürzlich gezeigt, dass sie für die Linsensteifigkeitwichtig sind 22,23.
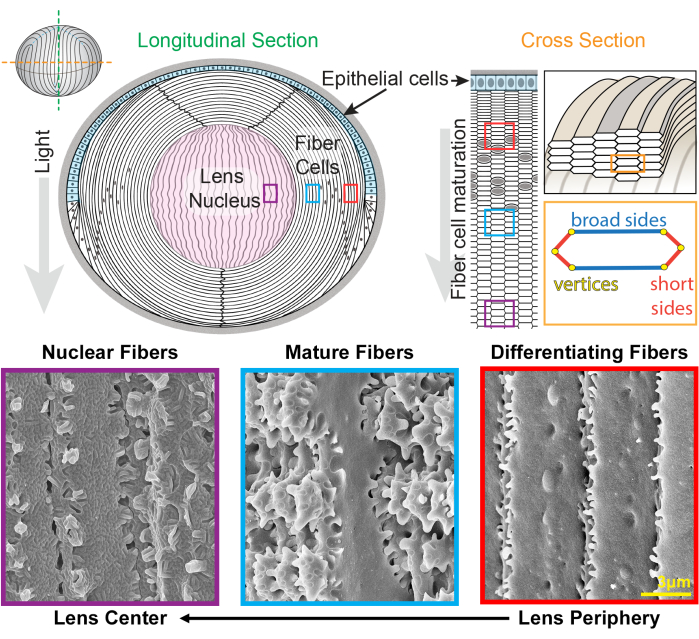
Abbildung 1: Linsenanatomiediagramme und repräsentative Rasterelektronenmikroskopie-Aufnahmen (REM) von Linsenfasern. Die Karikatur zeigt eine Längsansicht (von oben nach hinten von oben nach unten) der vorderen Monoschicht von Epithelzellen (hellblau schattiert) und einer Masse von Linsenfaserzellen (weiß). Das Zentrum der Linse (rosa schattiert) wird als Zellkern bezeichnet und besteht aus hochverdichteten Faserzellen. Auf der rechten Seite zeigt ein Querschnitts-Cartoon die längliche sechseckige Zellform von Linsenfasern, die in ein Wabenmuster gepackt sind. Faserzellen haben zwei breite Seiten und vier kurze Seiten. Repräsentative REM-Bilder entlang der Unterseite zeigen die komplexen Membranverflechtungen zwischen Linsenfaserzellen in verschiedenen Tiefen der Linse. Von rechts haben neu gebildete Linsenfasern an der Linsenperipherie kleine Ausstülpungen an den kurzen Seiten und Kugeln an der Breitseite (rote Kästchen). Während der Reifung entwickeln Linsenfasern große Paddeldomänen, die durch kleine Ausstülpungen entlang der kurzen Seiten (blaue Kästchen) verziert sind. Reife Faserzellen besitzen große Paddeldomänen, die durch kleine Ausstülpungen gekennzeichnet sind. Diese ineinandergreifenden Domänen sind wichtig für die biomechanischen Eigenschaften von Linsen. Faserzellen im Linsenkern haben weniger kleine Ausstülpungen entlang ihrer kurzen Seiten und haben komplexe Nut-Feder-Verflechtungen (violette Kästchen). Die Breitseiten der Zelle weisen eine globuläre Membranmorphologie auf. Die Karikatur wurde von22,32 modifiziert und nicht maßstabsgetreu gezeichnet. Maßstabsleiste = 3 μm. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
Die Linse wächst durch Hinzufügen von Schalen aus neuen Faserzellen, die über frühere Generationen von Fasern24,25 gelegt werden. Faserzellen haben eine längliche, sechseckige Querschnittsform mit zwei breiten Seiten und vier kurzen Seiten. Diese Zellen erstrecken sich vom vorderen zum hinteren Pol der Linse, und je nach Art können die Linsenfasern mehrere Millimeter lang sein. Um die Struktur dieser länglichen und dünnen Zellen zu unterstützen, schaffen spezielle Verflechtungen entlang der breiten und kurzen Seiten ineinandergreifende Strukturen, um die Linsenform und die biomechanischen Eigenschaften zu erhalten. Veränderungen der Zellmembranform während der Differenzierung und Reifung von Faserzellen wurden durch elektronenmikroskopische (EM) Studien umfassend dokumentiert 2,3,4,5,6,7,8,9,10,20,26,27,28,29 . Neu gebildete Faserzellen haben Kugeln und Pfannen entlang ihrer breiten Seiten mit sehr kleinen Vorsprüngen entlang ihrer kurzen Seiten, während reife Fasern ineinandergreifende Vorsprünge und Paddel entlang ihrer kurzen Seiten haben. Kernfasern weisen Nut-Feder-Verflechtungen und eine globuläre Membranmorphologie auf. Über die Proteine, die für diese komplexen ineinandergreifenden Membranen benötigt werden, ist wenig bekannt. Bisherige Studien zur Proteinlokalisierung in Faserzellen stützten sich auf Linsengewebeschnitte, die keine eindeutige Visualisierung der komplexen Zellarchitektur ermöglichen.
In dieser Arbeit wurde eine neuartige Methode entwickelt und perfektioniert, um einzelne und Bündel von Linsenfaserzellen zu fixieren, um die komplexe Morphologie zu erhalten und die Immunfärbung von Proteinen an der Zellmembran und im Zytoplasma zu ermöglichen. Diese Methode bewahrt die Zellmembranarchitektur, vergleichbar mit Daten aus EM-Studien, und ermöglicht die Färbung mit primären Antikörpern für spezifische Proteine. Wir haben zuvor immungefärbte kortikale Linsenfasern in Differenzierung und Reifung22,23. In diesem Protokoll gibt es auch eine neue Methode zur Färbung von Faserzellen aus dem Linsenkern. Dieses Protokoll öffnet die Tür zum Verständnis der Mechanismen für die Bildung und Veränderung der Membraninterdigitation während der Reifung von Faserzellen und der Verdichtung des Linsenkerns.
