The protocol described herein allows the collection of a minimum volume of 10-20 μl of insect hemolymph from individual caterpillars. Hemocytes collected using this method are free of cell clumping, melanization defects, tissue debris or other contaminants. Therefore hemocytes can be readily observed and counted under the microscope, and several insects can be observed within a few hosur. We observed that the majority of hemocytes in our samples consisted of plasmatocytes 3 (hemocytes spreading asymmetrically) (Figure 4). We are currently characterizing the different hemocyte types found in these caterpillars in detail (Stoepler et al., unpublished data). The method described here provides a quick and accurate means for counting total hemocyte numbers for individual caterpillars collected from field experiments.
Although hemocyte density can vary greatly between and within insect species 1, we found that hemocyte numbers for L. fasciola caterpillars range from 1.10 × 106 – 8.23 × 106 cells/ml of hemolymph (mean: 3.51 × 106; n = 31 caterpillars), and are in the range of previously reported values in other lepidopteran species 14.

Figure 1. Photograph of a penultimate instar Lithacodes fasciola (Lepidoptera: Limacodidae) caterpillar. Typical body length of these caterpillars ranges from 10-15 mm in penultimate to ultimate instars. Photo credit: John T. Lill.
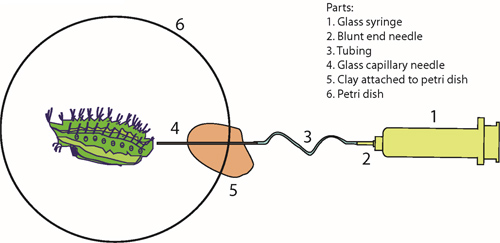
Figure 2. Injection needle setup. A glass syringe is connected to a capillary needle via a blunt end needle and plastic tubing (panels 1-4). A piece of plastic clay is used to rest the capillary needle (5). The caterpillar is placed in a Petri dish for injection with the collection solution (6).
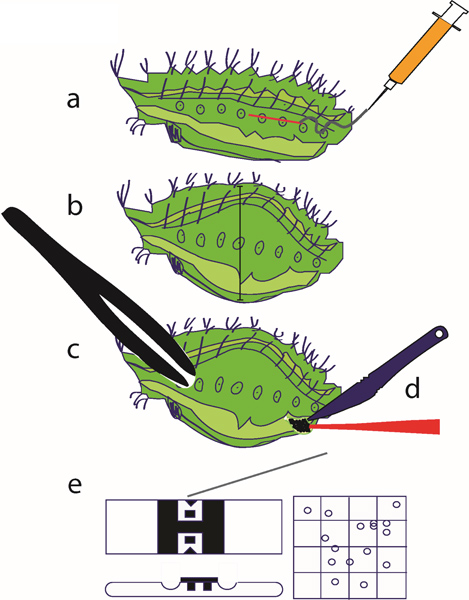
Figure 3. Hemocyte extraction method. (a) The chilled caterpillar is injected with the collection solution; (b) following injection, the caterpillar’s body will swell slightly, and the injected insect is allowed to rest on ice for 30 min; (c, d) a cut is made using a clean scalpel, the caterpillar is squeezed using a pair of plastic forceps to force the hemolymph out of the body through the incision, and hemocytes are collected with a pipette; (e) hemocytes are counted using a hemocytometer.
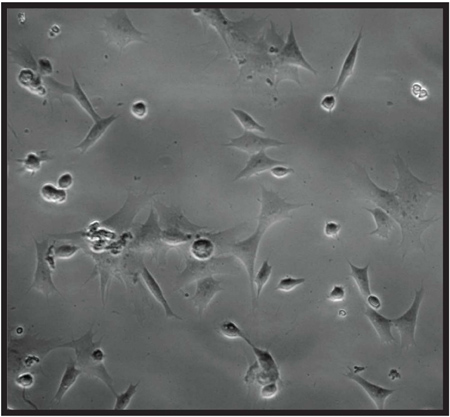
Figure 4. Microscopy picture showing extracted hemocytes from Euclea delphinii (Lepidoptera: Limacodidae) caterpillars (magnification: 20x).
