In the experiment described in Figure 2, EdU was administered 1.5 hr before irradiation and BrdU just after irradiation. Four types of cells were then distinguished in the cortical slices prepared at 1 or 4 hr post-irradiation, according to the incorporation of either EdU or BrdU, both or none (Figures 2A and 2B). Importantly, neither EdU nor BrdU incorporation changed the level of radiation-induced apoptosis (data not shown). Moreover, the staining methods allow only the detection of EDU and BrdU incorporated during DNA replication in S-Phase but do not have the sensitivity to detect DNA synthesis that is associated with repair even after irradiation. Indeed, i) neither EdU nor BrdU staining was observed at 1 hr post irradiation (PI) in cells of the cortical plate, where are located postmitotic neurons at G0 phase and ii) the number of EdU(+) and/or BrdU(+) was not increased in irradiated compared to unirradiated brains.
EdU(+)BrdU(+) nuclei
EdU(+)BrdU(+) nuclei correspond to NSPC that were in S phase before and after 0 hr PI. They were thus irradiated in S phase. As shown in Figures 3B and 3C their euchromatin is EdU(+) and may be also BrdU(+). Their chromocenters were either BrdU(+) or BrdU(-) depending on S phase completion at the time of sacrifice. Figure 2B shows that at 1 hr PI, EdU(+)BrdU(+) nuclei were localized in S phase bins (i.e. bins 4 to 10 , where NSPC are known to perform S phase31. Their distribution and numbers were not affected by irradiation. This suggests that radiation did not induce a complete block of DNA synthesis at this time.
Consistently with the basal to apical nuclear migration during G2 phase, most EdU(+)BrdU(+) nuclei of NSPC were found near the ventricle at 4 hr PI in unirradiated brains (Figure 2C). This did not occur in 2 Gy irradiated brains in which nuclei remained in S phase bins, many of them being apoptotic. Altogether this data showed that NSPC irradiated in S phase activated intra-S checkpoints in association with delayed INM and cell death.
EdU(+)BrdU(-) nuclei
EdU(+)BrdU(-) nuclei correspond to cells that were in S phase before and not after irradiation. Chromocenters of these nuclei are EdU(+), indicating that cells were in late S-phase when EdU was injected in mice (Figure 3A). Since G2/M phases of NSPC last 2 hr in the developing mouse brain, EdU(+)BrdU(-) NSPC were in G2 at 0 hr PI. Consistently, most EdU(+)BrdU(-) nuclei were found in the first bin near the ventricle in unirradiated brains at 1 hr PI. This is a consequence of INM of NSPC, after S phase, where many of them formed typical mitotic figures (Figure 2B). At 1 hr PI, the number of EdU(+)BrdU(-) nuclei was significantly reduced at the ventricular surface and no mitotic figure was detectable in irradiated brains. These data clearly show that cells irradiated in G2 activated the G2/M checkpoint within 1 hr PI. Furthermore, most of EdU(+)BrdU(-) of these nuclei were apoptotic at 4 hr PI demonstrating that NSPC are highly radiosensitive during G2 (Figure 2C).
EdU(-)BrdU(-) nuclei
Cells with an EdU(-)BrdU(-) nucleus correspond to either cells in G0 (principally immature neurons), or NSPC that remained in G1 from EdU incorporation to animal sacrifice (Figure 3E). Comparison between 1 hr and 4 hr in Figure 2C shows interkinetic nuclear migration making that nuclei that were close to the ventricle in G1 at 1 hr underwent their apical to basal migration at 4 hr, whereas nuclei that were in S phase went toward the ventricular surface.
As shown in Figure 2C many of these nuclei were apoptotic at 4 hr PI. The number of apoptotic EdU(-)BrdU(-) nuclei was maximal near the ventricular surface and decreases in upper bins. These data suggest therefore that radiation-induced apoptosis occurred in NSPC that have been irradiated during early G1 phase and/or post-mitotic migrating neurons. By contrast, none or very few apoptotic nuclei were found in the cortical plate. This is in accordance with the radiation resistance of neurons that have reached their final destination.
EdU(-)BrdU(+) nuclei
EdU(-)BrdU(+) nuclei correspond to cells that entered S phase after irradiation. Typically at 1 hr PI, they showed a diffuse BrdU labelling of euchromatin, but not of chromocenters. Thus they were in early S phase at that time (Figure 3D). At 1 hr PI the number and localization within S phase bins of EdU(-)BrdU(+) nuclei is similar between controls and irradiated brains. Thus S-phase entry during the 1st hr PI was not affected by radiation in the mouse developing brain (Figure 2B). Interestingly, comparison of 1 hr and 4 hr PI in irradiated brains shows a decline in the number of EdU(-)BrdU(+) nuclei and most of these nuclei were apoptotic at 4 hr PI. This suggests that cells that entered S phase after radiation activated intra-S checkpoints and underwent apoptosis thereafter (Figure 2C).

Figure 1. Analysis of Brain section Coronal section of the cerebral hemisphere of E14.5 mouse embryos stained with Dapi. EdU and BrdU staining as well as nuclear morphology are analyzed in a standard sector of the dorsomedial cerebral wall using a grid divided into 18 bins (or more) as described in the text. Scale bar: 20 µm. Please click here to view a larger version of this figure.
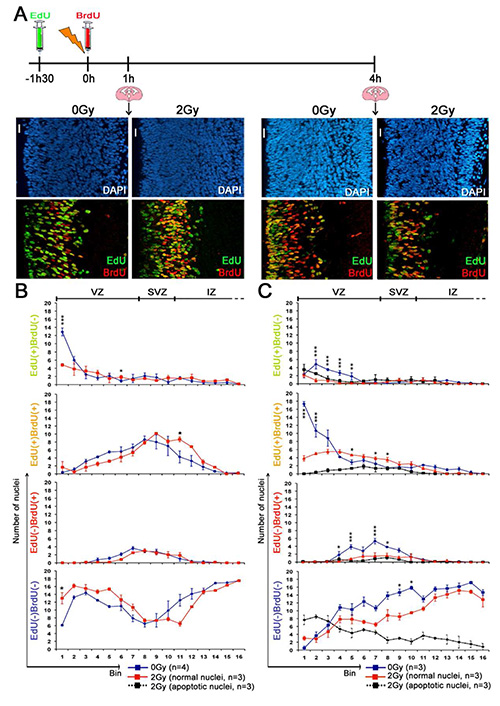
Figure 2. Cell cycle progression of irradiated neural stem and progenitor cell (Figure from Roque et al., Stem cells, 2012). (A) Schematic diagram of the experimental design and DAPI (blue), EdU (green), and BrdU (red) staining patterns found in coronal cortical slices at 1 and 4 hr PI (2 Gy). Scale bar = 20 μm. (B, C) Numbers/bin of EdU(+)BrdU(−), EdU(+)BrdU(+), EdU(−)BrdU(+), or EdU(−)BrdU(−) nuclei with a normal morphology in nonirradiated controls (blue) and with a normal (red) or apoptotic (pyknotic, black) morphology in irradiated (2 Gy) cortices at 1 hr (B) or 4 hr PI (C). No apoptotic nuclei were detected at 1 hr PI (B). Statistical analysis was performed using Bonferroni post hoc tests. Abbreviations: BrdU, 5-bromo-2′-deoxyuridine; DAPI, 4′-6-diamidino-2-phenylindole; EdU, 5-ethynyl-2′deoxyuridine; IZ, intermediate zone; SVZ, subventricular zone; VZ, ventricular zone. Please click here to view a larger version of this figure.
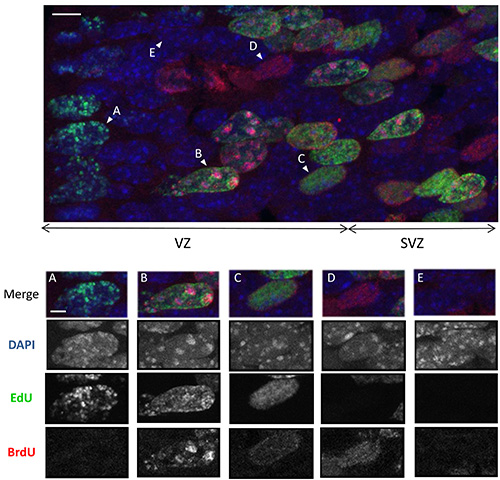
Figure 3. Differential EdU and BrdU staining of euchromatin and chromocenters Upper panel: Z-stack confocal images of EdU (green), BrdU (red) staining and DNA counterstained with DAPI (blue) allowing the detection of chromocenters as bright blue foci. Stainings were performed on an embryonic brain section collected at 1 hr PI in experiments where EdU was delivered 1.5 hr before irradiation and BrdU 0 hr PI (only the unirradiated control is shown) (Scale bar: 5 µm). Please click here to view a larger version of this figure.
Bottom panels show examples of the 5 different patterns of EdU and BrdU staining detected in coronal brain slices. Merge image and single channels are shown (Scale bar: 2 µm):
A) Nucleus with EdU(+) chromocenters and EdU(+) or EdU(-) euchromatin. EdU has been thus incorporated in late S phase during the replication of pericentric heterochromatin: the cells were in G2 at 0 hr PI.
B) Nucleus with EdU(+) euchromatin and BrdU(+) chomocenters: EdU has been incorporated in early S phase when most of euchromatin is replicated and BrdU has been incorporated in late S phase: the cells were in S phase at 0 hr PI.
C) Nucleus with EdU(+)BrdU(+) euchromatin and EdU(-)BrdU(-) chromocenters. EdU and BrdU have been incorporated in the first part of the S phase: The cells were in early S phase at 0 hr PI.
D) Nucleus with BrdU(+) euchromatin and EdU(-)BrdU(-) chromocenter. BrdU has been incorporated in the beginning of the S phase. The cells were in late G1 at 0 hr PI and entered S phase thereafter.
E) Nucleus with EdU(-)BrdU(-) euchromatin and chromocenters. The cells were in G0 (immature neurons) or in G1 phase and did not enter S phase during EdU and BrdU pulses.
| Solution | Incubation | Temperature | Pressure | Agitation | |
| 1 | Alcohol 70% | 30 min | 35 °C | on | on |
| 2 | alcohol 95% | 15 min | 35 °C | on | on |
| 3 | alcohol 95% | 30 min | 35 °C | on | on |
| 4 | alcohol 95% | 45 min | 35 °C | on | on |
| 5 | alcohol 100% | 15 min | 35 °C | on | on |
| 6 | alcohol 100% | 30 min | 35 °C | on | on |
| 7 | alcohol 100% | 1 hr | 35 °C | on | on |
| 8 | Toluene 100% | 30 min | 35 °C | on | on |
| 9 | Toluene 100% | 45 min | 35 °C | on | on |
| 10 | Toluene 100% | 1 hr | 35 °C | on | on |
| 11 | Paraffin | 30 min | 58 °C | on | on |
| 12 | Paraffin | 1 hr | 58 °C | on | on |
| 13 | Paraffin | 1 hr | 58 °C | on | on |
| 14 | Paraffin | 1.5 hr | 58 °C | on | on |
Table 1. Vacuum infiltration processor program for paraffin embedding.
| Reaction components for 1 slide | |
| 1X EdU reaction buffer | 128.64 µl |
| CuSO4 | 6 µl |
| EdU Alexa Fluor azide | 0.36 µl |
| 1X EdU Reaction buffer additive | 15 µl |
| Total volume | 150 µl |
Table 2. EdU reaction cocktails.
