The sharp, beveled injection needle is easily placed in the cardiac ventricle of the zebrafish larva when approached dorsally (Figure 3A). The heart continues pumping and blood flow is maintained despite the presence of the needle (Figures 3B and 3C). Careful injections do not disrupt the morphology of the ventricle or cardiac contractions (Figure 3D) despite injection of the morpholino into the heart (Figure 3E).
Within minutes, the morpholino-Endo-Porter solution distributes throughout the body via the vasculature (Figures 4A and 4B). The morpholino is then distributed from the vasculature into different tissues such as the finfold and brain for at least 12 hr or more as observed from the fluorescence (Figure 4C).
In this approach, we remove the distal tip of the finfold, the distal tip of the spinal cord, distal trunk muscle, the distal notochord and pigment cells (Figures 4D and 4H); all of which are difficult to specifically target by direct injection due to compactness (muscle), small size (spinal cord and notochord) and thinness (finfold), but appear to incorporate the morpholino after serial ventricular injections, which is seen by fluorescence within these tissues (Figure 4E) compared to uninjected animals (Figures 4F and 4G). Thus, this method relatively easily promotes the delivery of morpholino to tissues that are difficult to individually target, and it permits the assessment of gene function in several tissues in the regenerating fin at once. To assess the importance of specific genes involved in regeneration, one surgically partially amputates the larval caudal fin (Figure 4H), resecting all the tissues that have incorporated the morpholino (Figure 4I). The larval finfold regenerates its structure within a few days post amputation (dpa) (Figure 4J). Morpholino targeting a gene required for regeneration (unpublished data) results in a perturbed regeneration response (Figure 4K) compared to uninjected (Figure 4J) and mismatch morpholino controls (Figure 4L). The total effect of these morpholino experiments on regenerative outgrowth can be measured using standard morphometric tools in the publically available Fiji programs12 by tracing the regenerated tissues and comparing the areas (Figures 4J-4L). Thus, this gene-targeting method provides a quick and relatively easy assay to test the importance of genes in the regeneration process.

Figure 1. Preparation of glass capillary needles for injection. A) Glass capillary tube before pulling (above) and pulled needle (below). B) Needle pulling apparatus. C) Lathe for beveling and sharpening the glass needle. The needle is viewed through the binoculars. D) The lathe is a wetted rubber spinning disc. The needle is slowly lowered onto the disc. E) Sharpened needle must have a short bevel that is no longer (20 µm) than is wide (20 µm) to minimize the space needed to insert the entire end of the needle into the larval cardiac ventricle. F) Higher magnification image showing the tip of the needle. G) Schematic view of the needle tip showing the shape of the bevel after sharpening.

Figure 2. Injection apparatus set up. A) Complete apparatus for injection of morpholino into zebrafish larva. B) Stand to control the placement of the needle during injections. C) The pico pump that regulates pneumatic pressure for injections. D) Form press used to create agarose mold. E) Agarose mold used to stabilize larva for injection.
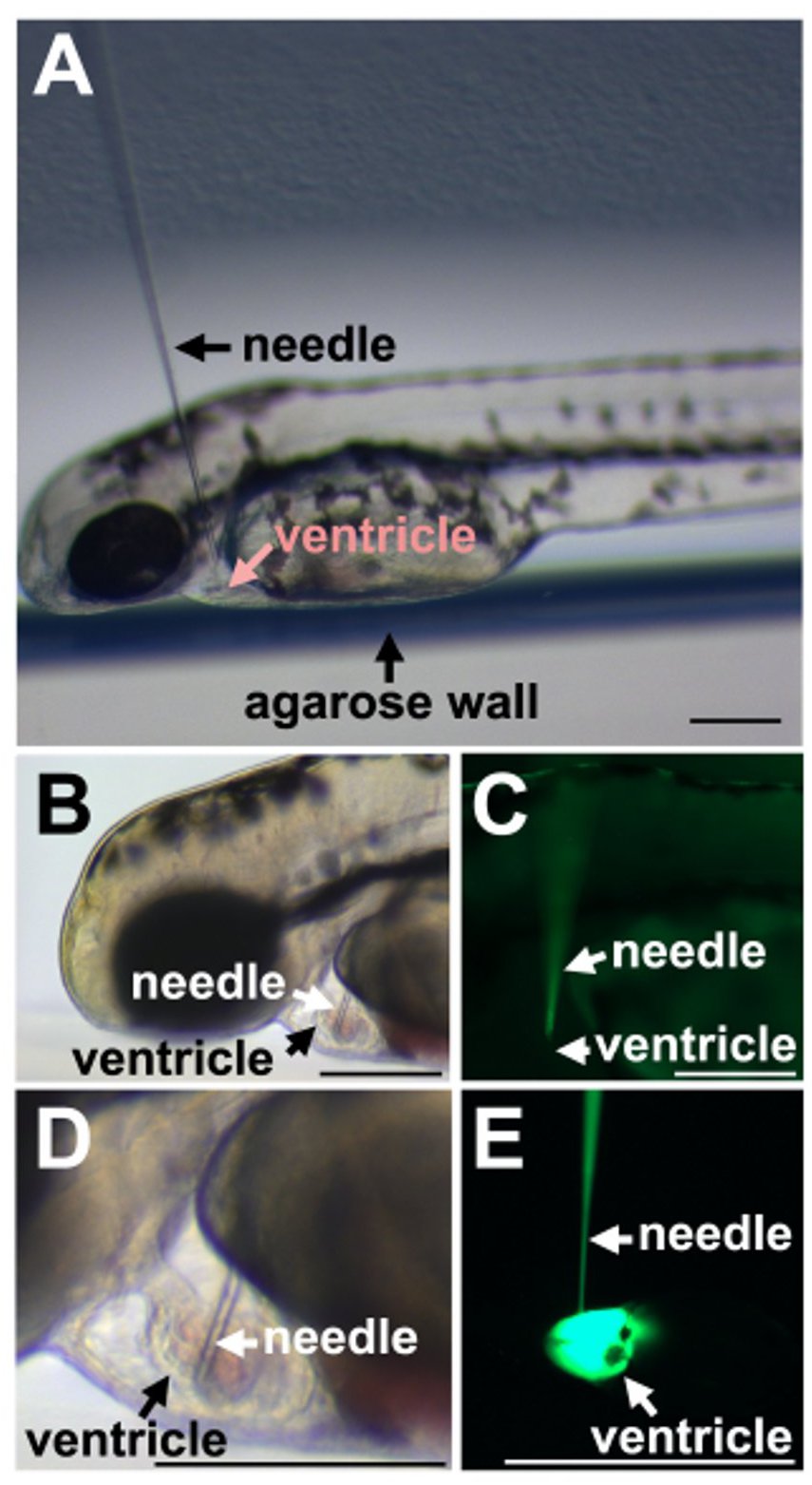
Figure 3. Injection of fish larva. A) Placement of the needle in relation to the zebrafish larva. B) Insertion into cardiac ventricle. C) Fluorescence of the morpholino in the needle but absent in the ventricle before injection. D) Brightfield image shows the size relationship between the glass needle and the larval ventricle. E) Injection of the fluorescent morpholino into the heart ventricle. Scale bars equal 100 µm.
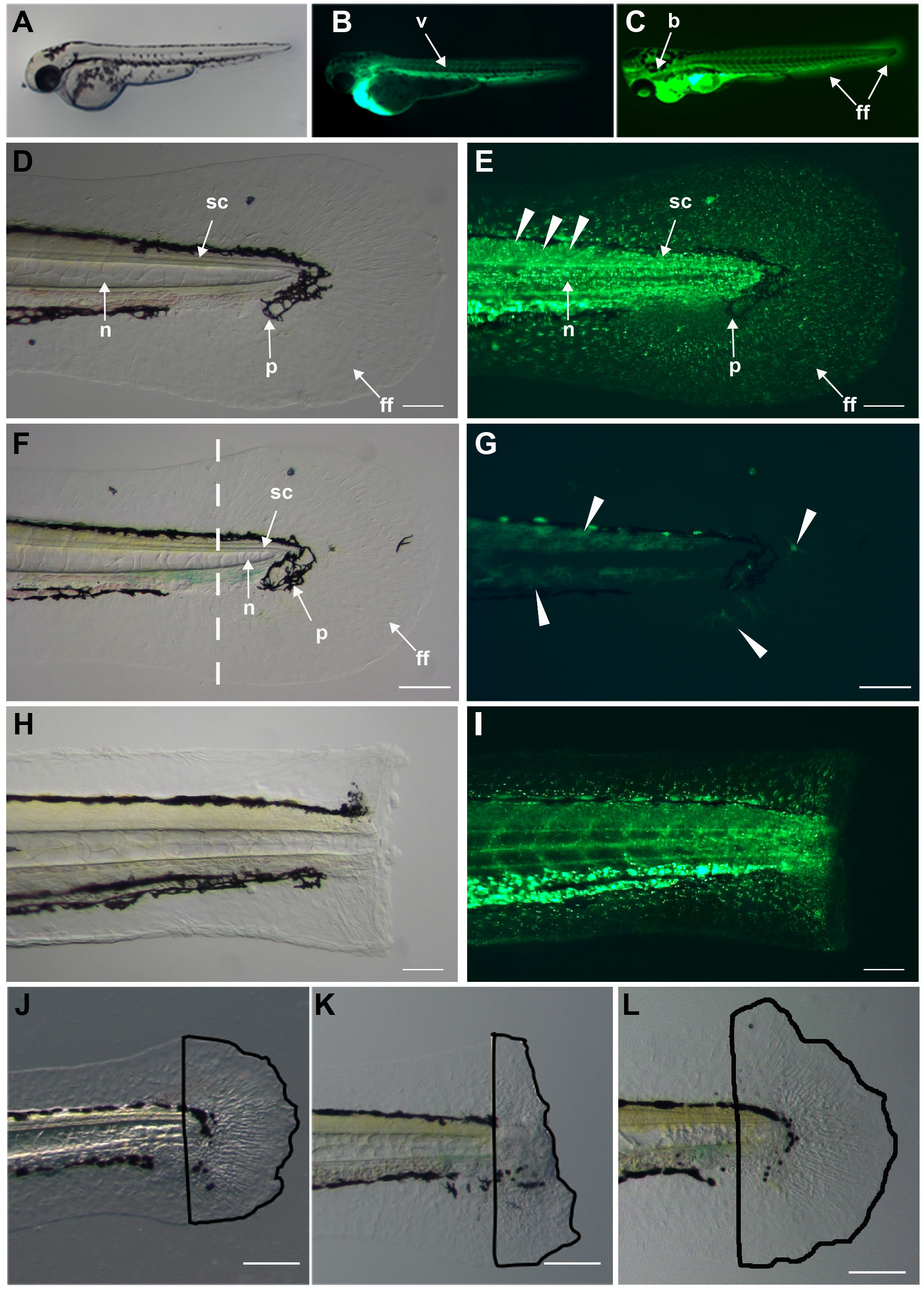
Figure 4. Dispersion of morpholino throughout fish. A) Zebrafish 3 day old larva. B) Fluorescence of fluorescein-tagged morpholino throughout the body vasculature (v) 5 min post ventricular injection. C) Dispersion of the morpholino throughout the animal including finfold (ff), brain (b) in older larva after injections of morpholino every 12 hr. D) Tissues in the larval caudal fin and trunk: notochord (n), spinal cord (sc), pigment (p), and finfold (ff). E) Dispersed morpholino several minutes after injection into larval trunk tissues including the finfold. F) Brightfield image of uninjected larval trunk and caudal fin. Dashed line indicates the prospective amputation plane. G) Uninjected fin imaged with the GFP filter showing autofluorescence in green (arrowheads). The dendritic shape of the fluorescing structures suggests that these are pigment cells. H) Brightfield image of a surgically amputated larval caudal fin through the notochord, muscle and spinal cord. I) Fluorescent image showing morpholino distribution in the stump tissues. Morpholino incorporation into muscle, spinal cord, notochord and finfold. J) Regenerated caudal finfold structures of wild-type larva. Black line traces the regenerated tissues for quantification analysis. K) Morpholino-mediated inhibition of regeneration. Black line tracing clearly highlights the reduced regeneration response after morpholino knockdown. L) Regenerated caudal finfold of larva injected with a mismatch control morpholino showing a similar regeneration response as in uninjected larva (J). Scale bars equal 100 µm.
