For the first set of experiment, we microinjected 200 nL of AAV9.hSynapsin.HI.eGFP-Cre.WPRE.SV40 viruses (AAV-eGFP-Cre, 1/10 dilution in Dulbecco's phosphate buffered saline) that express the Cre DNA recombinase fused with GFP into P2 striatum of Ai14 mice. The Ai14 mice express tdTomato reporter gene upon Cre-mediated deletion of loxP-flanked (floxed) STOP cassette (Figure 2F). The brains were harvested at P14 for immunostaining of GFP and tdTomato. Many AAV transduced GFP-positive cells were present throughout the striatum (Figure 2A, C), indicating an extensive infection of striatal cells by AAV-eGFP-Cre viruses. Similar extensive expression of tdTomato was also found in the striatum (Figure 2B, C). Upon microscopic examination at high magnification, we found that nearly all GFP-positive striatal cells co-expressed the tdTomato reporter gene (Figure 2A'-C'). Moreover, tdTomato signals were detected in presumably axon terminals in the globus pallidus (Figure 2D) and the substantia nigra pars reticulata (Figure 2E), the target regions of striatonigral and striatopallidal projection neurons13. These results suggest a successful Cre-loxP mediated DNA recombination induced by AAV-mediated expression of Cre activity in neonatal striatal neurons.
For the second set of experiment, we microinjected 50 nL of AAV-eGFP-Cre viruses into the striatum of Foxp2fl/fl mice carrying loxP-flanked Foxp2 alleles at P2 (Figure 3A-C, F), and harvested the brain at P9. No Foxp2+;GFP+ double-labeled cells were found in the striatum, i.e., Foxp2 protein was absent in GFP-positive cells (Figure 3A'-C'). In control experiments, Foxp2+;GFP+ double-labeled cells were present in the striatum of Foxp2fl/fl mice with AAV-eGFP control viruses (Figure 3D) and in the striatum of heterozygous Foxp2fl/+ mice with AAV-eGFP-Cre viruses (Figure 3E). These results indicate that Foxp2 gene is specifically deleted in AAV-eGFP-Cre transduced cells of neonatal striatum.
Taken together the results of AAV-eGFP-Cre;Ai14 and AAV-eGFP-Cre;Foxp2fl/fl mice, the technique of stereotaxic neonatal brain surgery that we developed is amenable for conditionally delete genes in specific regions of neonatal mouse brains.

Figure 1. Stereotaxic apparatus and homemade head-fixed holder for neonatal mice. (A) The stereotaxic injection system consists of the following devices: i. syringe pump (controller); ii. syringe pump (drive unit); iii. microliter syringe (10 µL); iv. PE20 tubing adaptor; v. PE10 tube; vi. 30G injection needle; vii. stereotaxic apparatus; viii. a pipette tips box converted platform for neonatal pups; xi: crushed ice. (B) The height of the tissue embedding cassette is about 7 mm. The button of 1.5 mL centrifuge tube is about 15 mm long. (C) The mouse head is secured in the homemade head-fixed holder. The arrow indicates the landmark lambda in the head of neonatal mice. Please click here to view a larger version of this figure.
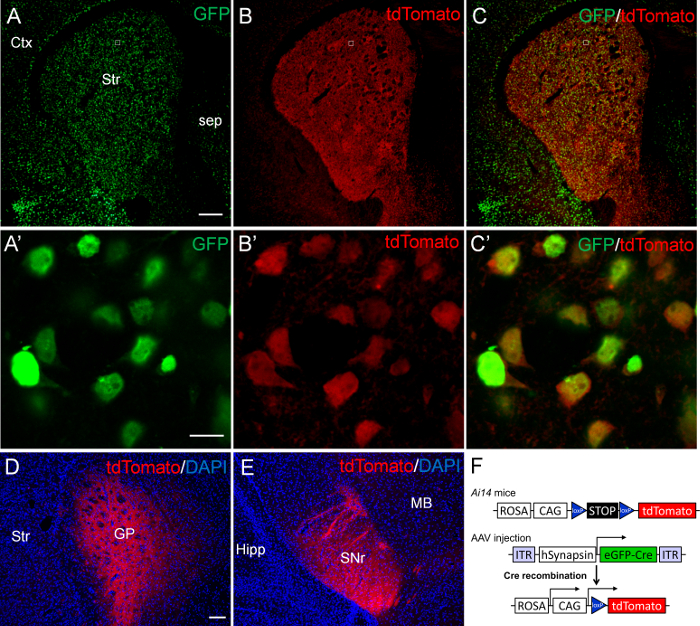
Figure 2. Expression of tdTomato in the striatum of P14 Ai14 mice after AAV-eGFP-Cre mediated Cre-loxP DNA recombination. AAV-eGFP-Cre viruses were sterotaxically injected into P2 striatum of Ai14 mice. The brain was analyzed at P14. (A) Many GFP-positive cells are present throughout the striatum (Str). (B) Many tdTomato-positive cells are also present in the striatum. (C) The merged images show an extensive co-localization of GFP and tdTomato in the striatum. Confocal images of the boxed regions in A-C are shown at high magnification in A'-C', respectively. All GFP-positive cells (green) co-express tdTomato (red) in the striatum. (D, E) The tdTomato signals are detected in the target regions of striatal projection neurons, including the globus pallidus (GP; D) and the substantia nigra pars reticulata (SNr; E). (F) Schematic drawings of AAV-eGFP-Cre and Ai14 transgenic constructs. Ctx: cortex; Sep: septum; Hipp: Hippocampus; MB: midbrain. Scale bar in A (for A-C), 200 µm; A' (for A'-C'), 10 µm; D (for D and E), 100 µm. Please click here to view a larger version of this figure.
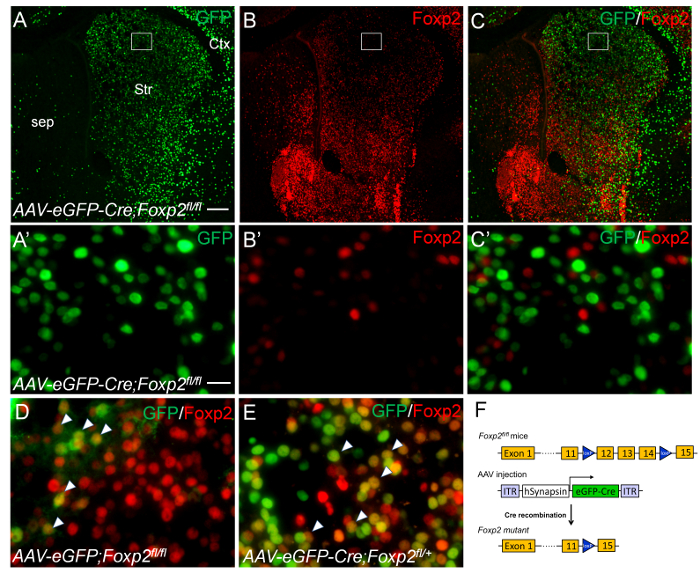
Figure 3. Conditional deletion of Foxp2 gene in the neonatal striatum by intrastriatal injecions of AAV-eGFP-Cre viruses. Intrastriatal injections of AAV-eGFP-Cre viruses were performed in P2 Foxp2fl/fl (A-C') and Foxp2fl/+ (E) mice. The brains were analyzed at P9. (A-C) GFP-positive cells (A, C, green) lack Foxp2 immunoreactivity (B, C, red) in the striatum of Foxp2fl/fl mice with AAV-eGFP-Cre viruses. The boxed regions in A-C are shown at high magnification in A'-C', respectively. (D) Striatal cells co-express GFP and Foxp2 are present in the striatum that was injected with AAV-eGFP control viruses. (E) GFP-positive cells co-express Foxp2 (arrowheads) are present in the striatum of Foxp2fl/+ mice with AAV-eGFP-Cre viruses. (F) Schematic drawings of AAV-eGFP-Cre virus and Foxp2fl/fl transgenic mice. Scale bar in A (for A-C), 200 µm; A' (for A'-C', D and E), 20 µm. Please click here to view a larger version of this figure.
