Placentae are collected from healthy term, not in labor women undergoing elective cesarean sections. The Collection, processing, and disposable of human samples adhere to the guidelines of the Mount Sinai Hospital Ethics board. A written consent is obtained from each patient. This study is approved by the Research Ethics Board at Mount Sinai Hospital.
1. Preparations
NOTE: All steps must be conducted under a fume hood and all surgical equipment must be sterilized via autoclave prior to placement in the fume hood. All other materials (bottles, 50 mL tubes, etc.) must be sterilized with 70% ethanol solution. Always wear personal protective equipment at all times when working with biohazardous waste (lab coat, gloves, long hair tied back, etc.).
- Enzymatic digestion solution (Final solution: 200 mL)
- Pipette 180 mL of HBSS-/- into a 500 mL beaker, add sterile stirring magnet and place on stirring plate at room temperature.
- Weigh out and add the following to the HBSS-/- solution slowly and sequentially: 20 mL of FBS, 400 mg Collagenase 2 ([final] = 2 mg/mL), 20 mg Soy bean trypsin inhibitor ([final] = 0.1mg/mL), 30 mg DNase 1 ([final] = 0.15 mg/mL), and 200 mg BSA ([final] = 1 mg/mL)
- Set the stirring to medium speed, and allow the mixture to stir for 10-20 min (cover with tin foil or paraffin film while stirring).
- Pour the mixed solution into a 250 mL glass bottle using a plastic funnel and place in the fume hood.
- Pass the solution through a plastic top filtration unit (0.22 μm membrane filter, 500 mL), aliquot 20 mL into 50 mL tubes (10 tubes total) and store at -20 °C until use.
- RPMI 1640 Media (2% washing solution and 10% complete growth medium)
- Combine RPMI 1640 and FBS in a glass bottle in the fume hood.
- For 2% FBS solution, combine 490 mL of RPMI 1640 and 10 mL FBS.
- For 10% FBS solution, combine 450 mL RPMI 1640 and 50 mL FBS.
- Pipette 500 μL of normocin into the 500 mL glass bottle from the previous step containing RPMI media and FBS (50 mg/mL stock, 0.05 mg/mL working concentration).
- Pass media through a 0.22 μm membrane filter and store in a 500 mL glass bottle at 4 °C until use.
- 30 min prior to starting the experiment, place a 20 mL aliquot of frozen enzymatic digestion solution in a 37 °C bead bath, place the 2% FBS and the 10% FBS RPMI 1640 media in a 37 °C bead bath, turn on the rocking water bath and set to 37 °C, 2 g, and set a temperature controlled centrifuge to 4 °C.
2. Collection of Decidual Tissue from Term Placental Membranes
- Place bottles with HBSS+/+, HBSS-/- and five 50 mL tubes in a rack under the fume hood. Pipette 25 mL of HBSS+/+ in one tube.
- Using gloved hands, take the term placenta out from the container used for transporting it from the operating room theatre. Place the placenta on the diaper pad (maternal side up) and spread the fetal membranes out using scissors and forceps.
- Lay overlapping diaper pads on the fume hood.
- Use a separate diaper to flip the placenta so the maternal side is face up.
- Find the point of membrane rupture and make incisions to allow the membrane to unfold and lay flat on the fumehood surface.
NOTE: Once the membrane is pulled back from the placenta, the decidua will be face up resting on the chorion layer, with the amnion layer at the back.
- Using a plastic cell scraper, carefully scrape the decidual tissue off the chorion and place in the 50 mL tube containing 25 mL HBSS+/+.
- Scrape the membrane with moderate pressure. Do not apply too much pressure as it may result in chorion contamination. Collect small blood clots as well, as these contain some decidual cells.
Caution: After decidual collection, placenta must be packaged in a biohazard plastic bin and frozen in a -20 °C freezer prior to appropriate disposal (according to your institutional rules). Bloody diaper pads must be wrapped in a seal-tight plastic bag and disposed of in a biohazard safety box.
3. Washing and Enzymatic Digestion of the Decidual Tissue
- Wash the collected tissues by gently shaking the 50 mL tube by hand and passing it through a 250 μm metal sieve (250 μm, size 60 mesh) resting over a sterile specimen (urine) container.
- Repeat the wash twice with HBSS+/+ and twice with HBSS-/- in fresh tubes for each wash. (final washes with HBSS-/- allows for the removal of calcium and magnesium present in HBSS+/+, which would otherwise interfere with the following enzymatic digestion process).
NOTE. During the washing steps, chorion tissue contamination will be apparent as the decidual tissue is light pink in color and amorphous, while the chorion is white, dense, and stringy. Therefore, chorion contamination can be easily removed with forceps. - Optionally, if the decidual tissue is thick, transfer it to a sterile 10 cm diameter petri dish and proceed to mince of the tissue with two opposing scalpels in the dish.
- Place the washed tissue in sterile 50 mL tube containing 100 mg of tissue/mL of enzymatic digestion solution (see Preparations).
- As a reference point, ensure that the level of decidual tissue reaches the 5-10 mL mark on the 50 mL tube.
- Use approximately 20 mL of enzymatic digestion solution (Prepared in step 1.1) to digest the total decidua collected from a whole term fetal membrane.
- Seal the cap of the tube with the paraffin film (seal the tube tightly and wrap paraffin film around the cap and top of the tube) under the culture hood and incubate decidual tissue at 37 °C for 20 min in a shaking water bath (145 rpm, 2 g).
- After incubation, remove the paraffin film and sterilize the surface of the tube containing digested tissue with 70% ethanol and bring under the fume hood. Shake the tube briefly by hand.
- Collect cell suspension through the metal sieve (250 μm, size 60 mesh) into a new sterile specimen container. Dilute with an equal volume (20 mL) of RPMI + 10% FBS containing 0.1% normocin to stop enzymatic reaction. Proceed directly to centrifugation step 4.1.
- If necessary, place the remaining undigested tissue back into a new 50 mL tube with 20 mL of fresh enzymatic digestion solution and repeat digestion (20 min, 37 °C, shaking water bath).
- If a second digestion is necessary, place the first tube with cell suspension on ice (cover with paraffin film to keep sterile). Repeat steps 3.5-3.6 and combine the two cells suspensions.
4. Generating a Single Cell Suspension
- Centrifuge the cell suspension (420 g, 4 °C, 11 min).
- Remove the supernatant and resuspend the cells in 40 mL of RPMI + 2% FBS containing 0.1% normocin ("wash buffer").
NOTE: Cell pellet will be loose and gelatinous due to red blood cell contamination, aspirate supernatant with caution. Removal of the upper phase with a manual pipette may be required. - Repeat the centrifugation at 420 g at 4 °C for 11 min.
- Carefully remove the supernatant (As mentioned in Note Step 4.2) and resuspend the cell pellet in 5 mL of wash buffer and add 35 mL of erythrocyte lysis buffer in the same tube.
NOTE: If the pellet is large or very bloody, it can be split into two tubes for the erythrocyte lysis step by adding 10 mL of wash buffer and dividing equally into two tubes and then adding 35 mL of Erythrocyte lysis buffer to each tube. - Incubate on ice for 20 min. Briefly vortex tube(s) at the beginning and the end of the incubation to lyse the red blood cells.
- Centrifuge at 420 g for 11 mins at 4 °C.
- Carefully, remove the supernatant and resuspend the pellet in 40 mL of wash buffer.
- Pass the cells through a 70 μm nylon filter to remove cell clumps.
- Centrifuge at 420 g for 11 min at 4 °C.
- Remove the supernatant and resuspend the cell pellet in 10 mL of complete medium (RPMI 10% + FBS containing 0.1% normocin).
- Count cells using the trypan blue dye-exclusion hemocytometer procedure as described below:
- Under the culture hood gently pipette cell suspension up and down 3x in order to thoroughly mix before combining with trypan blue. In a 1.5 mL tube prepare cell suspension, diluted 1:2 (combine 20 μL of trypan blue solution and 20 μL of decidual cell suspension). Carefully mix trypan blue-cell solution by pipetting up and down a couple of times (this solution is not sterile).
- Place the hemocytometer on the stage of the microscope with glass coverslip on top.
NOTE: Hemocytometer is a microscope slide with grids on it to give nine large squares divided by triple lines. Each large square has an area of 1 mm2, and the depth of fluid in the chamber is 0.1 mm. Therefore, the volume of fluid that can fill each large square is 1 mm*1 mm*0.1 mm= 0.1 mm3= 10-4 mL. - Slowly add 10 μL of the trypan blue-cell mixture into the groove of the hemocytometer, allowing capillary action to disperse the cell mixture over the entire slide (stop before mixture fills well).
- View the cells under a microscope (at 10X magnification) and count all white/green cells that exclude trypan blue (these are the viable cells) in the four large outer squares of the hemocytometer. When counting cells that touch the line, count only those that touch the right and upper lines but not those touching the left and bottom lines. Do not count dark blue cells; blue color indicates that the cell is dead as trypan blue dye can easily penetrate through the plasma membrane into the cytoplasm.
- To calculate the number of viable cells in 1 mL of cell suspension (X):
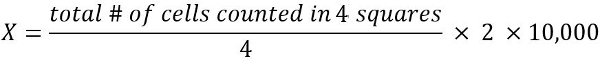
2 = dilution factor
10,000 = conversion factor (1 mL = 1 cm3 = 10,000*0.1 mm3)
- Dilute decidual cells to a desirable final concentration in RPMI + 10% FBS (growth medium).
NOTE: For tissue culture plating, pipette 2*106 cells/well into a plastic 6-well plate, 10*106 cells into a 10-cm plastic plate or 75,000 cells to a glass coverslip.
To validate the efficiency and viability of the isolated cells, they were characterized by two methods: flow cytometry and immunocytochemistry (ICC). 4 cell populations were targeted; decidualized stromal cells were detected by the anti-vimentin antibody, pan-leukocyte marker CD45 was used to identify decidual immune cells, cytokeratin was used to detect epithelial/endothelial cells and finally, cytokeratin 7 was used to detect any potential trophoblast (chorion or placental) contamination.
For characterization by multicolor imaging flow cytometry, freshly isolated decidual cells were stained with a fixable dye to determine cellular viability. The cells were permeabilized and fixed, followed by staining with fluorochrome-conjugated mouse monoclonal antibodies targeting vimentin-APC, CD45-APC-H7, cytokeratin-FITC and cytokeratin 7-PerCP. Antibody information is listed in Table of Materials. Fixed and stained cells were stored at 4 °C overnight in staining buffer to prevent dissociation of tandem dye and run the following day on the Image stream MK2 imaging flow cytometer. The analysis was performed using a software. Cell debris and aggregates were excluded by serial gating using three sequential feature/mask combinations (Figure 1A-B). Fluorescence Minus One (FMO) gating controls were used to identify positive decidual cell populations and single stain cells were used for compensation (Figure 2). Final dot plots demonstrate the positively stained cell population of the four markers (Figure 2) and the cellular viability of 80% (Figure 1C). Representative images of positively stained cells can be seen in Figure 1D. Using this method, we found that the population mainly comprised of stromal cells (55-60%), leukocytes (35%), epithelial (1%) or trophoblast (0.01%) cells (Figure 1E). The experiment was repeated three times. These results verify the purity (absence of trophoblast contamination from the chorion) and high viability of primary human decidual cells collected and isolated from the fetal membranes of term placentae.
In a separate ICC experiment, freshly isolated decidual cells were seeded on glass coverslips (75,000 cells/coverslip), allowed to attach and remained in culture for 48 hours. Media was changed after 24 hours to remove non-attached dead cells. The cells were then fixed with 50% acetone-50% methanol, permeabilized with 0.02% non-ionic surfactant and unspecific binding was blocked with blocking solution. The cells were then stained with mouse monoclonal antibodies: anti-CD45 (pan-leukocyte marker), anti-cytokeratin (epithelial cell marker) and anti-cytokeratin 7 (trophoblast marker), and goat polyclonal anti-vimentin antibody (stromal cell marker). Donkey anti-goat and donkey anti-mouse were used as secondary antibodies. DAPI was used to stain the nuclei. Mouse IgG, Goat IgG and secondary antibodies alone were used as negative and specificity controls. Images were taken at 20X magnification on a spinning disc confocal microscope. Antibodies used are listed in Table of Materials and representative images are presented in Figure 3. Our visual observation indicates that majority of attached primary decidual cells are vimentin-positive stromal cells (approx. 95%), with a small number of leukocytes (approx. 1-2%), epithelial (approx. 1%) and trophoblast cell populations (single cells were detected accounting for <0.01% of all). The major difference between the flow cytometric and ICC results is the decrease in CD45 positive leukocyte population detected by flow cytometry. This discrepancy can be explained by the methods in which the two experiments were conducted: in the flow cytometry analysis, isolated decidual cells were immediately processed and stained, meaning the leukocyte population was included in analysis. With ICC, the total decidual cell suspension was plated and cultured for 48 hours in media that was mostly supporting the attachment of stromal and epithelial cells, but not immune cells. After 24 h the immune cells that were not attached were removed during media change, accounting for the difference in results presented by these two methods. We concluded that for studying leukocyte sub-populations in human decidua from complicated pregnancies special precaution should be used to avoid accidental elimination of the leukocyte population.
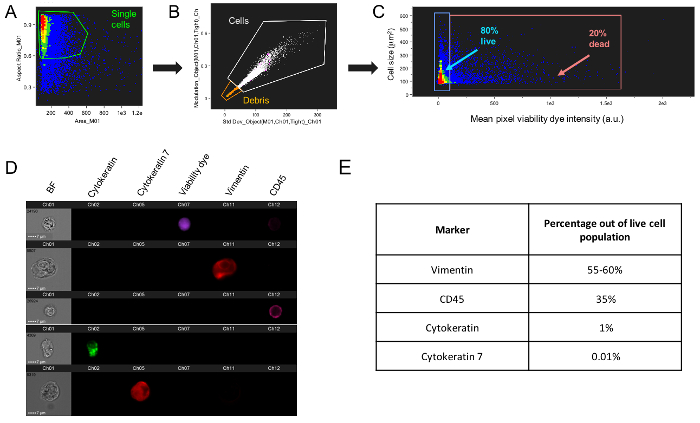
Figure 1: Gating strategy for decidual cell population discrimination. (A) Freshly isolated fixed decidual cells were initially gated to exclude cell doublets, and (B) subsequently gated using three sequential feature/mask combinations to exclude cell debris. (C) Live cells (pink gate, 80% of total population) were differentiated from dead cells (blue gate, 20% of total population) using a fixable viability dye. (D) Representative images of decidual cells stained with viability marker and four fluorescent-conjugated antibodies (Ab) (vimentin-APC, CD45-APC-H7, cytokeratin-FITC and cytokeratin 7-PerCP). Dotted scale bar equals 7 μm. (E) Once a debris-free, live cell population was determined, these markers were used to determine the percentages of each cell type within the decidual cell population. For Figures 1B and 1C, each dot represents a single decidual cell. For Figures 1A and 1C, the color range of the dot plots depicts cell density, with red signifying highly dense cells and blue less dense of cells. Data presented here is representative of 3 biological replicates. Please click here to view a larger version of this figure.
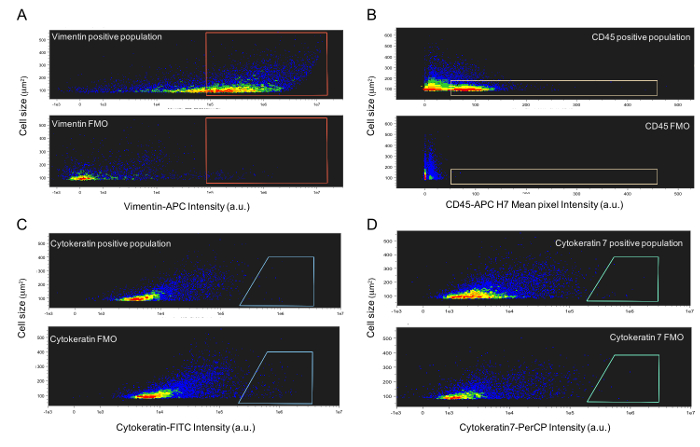
Figure 2: Representativeflow cytometric analysis of primary human decidual cells with FMO gating plots. Representative color dot plots of cells isolated from term decidua and stained with (A) vimentin-APC (stromal cell marker), (B) CD45-APC-H7 (leukocyte marker), (C) cytokeratin-FITC (epithelial cell marker) and (D) cytokeratin 7-PerCP (trophoblast marker) alongside each respective FMO control. Fluorescent-Minus-One (FMO) controls, tubes which contain the full antibody cocktail minus the marker in question and prepared identically to the fully stained experimental sample, were used during flow cytometry data analysis to determine true positive decidual cell populations. FMO controls show the relative background and allow for accurate gating of true positive signals. Data presented here is representative of 3 biological replicates. Please click here to view a larger version of this figure.
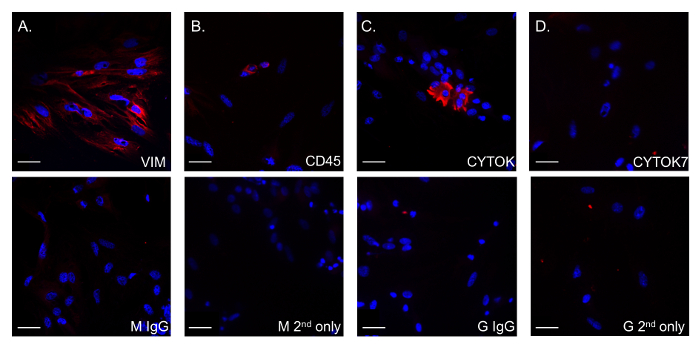
Figure 3: Representative Immunocytochemical analysis of primary human decidual cells. Primary human decidual cells were grown on glass coverslips for 48 h, with media changes after 24 h. Cells were fixed with acetone-methanol and stained with monoclonal mouse anti-CD45 (leukocyte marker), anti-cytokeratin (epithelial cell marker) and anti-cytokeratin 7 (trophoblast marker) antibodies, and goat polyclonal anti-vimentin antibody (stromal cell marker). Representative images show (A) vimentin-positive fibroblasts (secondary Abs: donkey anti-goat), (B) CD45+ leukocytes (secondary Abs: donkey anti-mouse), (C) cytokeratin positive epithelial cells (secondary Abs: donkey anti-mouse) and (D) cytokeratin 7-positive trophoblast cells (secondary Abs: donkey anti-mouse) were detected (red staining). DAPI staining was used to dye the nuclei of cells (blue staining). Non-specific mouse and goat IgG (M IgG and G IgG) were used as negative controls, omitting primary Abs were used as secondary antibody controls (M 2nd only and G 2nd only). Images are taken at 20X magnification on a DMI spinning disc confocal microscope. Scale bars = 32 μm. Please click here to view a larger version of this figure.
