To visualize the interaction between Qdot-labeled ORC and the ARS, we first constructed the λ-ARS317 DNA substrate. A DNA fragment containing ARS317 was integrated into XhoI site (33.5 kb) of native λ DNA by homologous recombination (Figure 1A). The recombination product was packaged using extracts and the packaged phage particles were cultured on LB plates (Figure 1B). The positive phage plaque was screened by PCR and confirmed by sequencing (Figure 1C). λ-ARS317 DNA was purified from the liquid lysates (Figure 1D).
Single-molecule imaging assays were carried out on the objective-type TIRFM set-up (Figure 2). By labeling the biotinylated ORC with a nearly 1:1 molar ratio streptavidin-coated Qdots rapidly, Qdot-labeled ORC was pumped into the λ-ARS317 tethered flow cell. DNA was stained using SYTOX Orange. The signals of Qdot-labeled ORC and SYTOX-Orange-stained DNA were imaged concurrently using TIRFM (Figure 3A-3C). To determine the distribution of ORC on λ-ARS317, the imaging stack was separated into two stacks: one stack of DNA signal and the other stack of ORC signal as described in step 6.1 (Figure 3B). Based on the formula, Position (kb) = (LORC/LDNA) × 50kb (Figure 3D), 273 binding positions of ORC-Qdot705 molecules on 168 DNA substrates were quantitively analyzed. The data showed that ORC-Qdot705 specially bindson the ARS317 inserted site with an obviously high abundance (Figure 3E). Meanwhile, ORC-Qdot705 also binds on the AT-rich areas located in middle and the free end of λ DNA, which is consistent with the results of the previous study10.
The high-quality image acquisition depends on the molar ratio of protein and Qdots in the labeling assay. Excessive Qdots may be good for the protein labeling efficiency, but they can create background noise. As shown in Figure 4, while labeling the biotinylated ORC with streptavidin-coated Qdots at 1:3 molar ratio, the background noise increased. Thus, it is critical to choose the appropriate molar ratio of biotinylated proteins to streptavidin-coated Qdots.
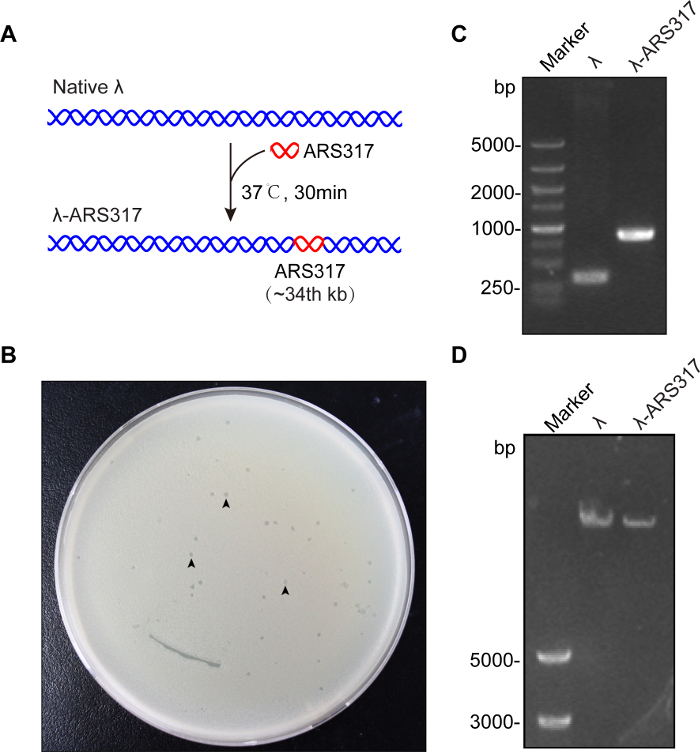
Figure 1: Preparation of λ-ARS317 DNA substrate. (A) Illustration of λ-ARS317 construction. A DNA fragment bearing ARS317 was integrated into λ DNA by homologous recombination. (B) Plaques on LB solid medium. Three of them were marked using black arrows. (C) A λ-ARS317 plaque was identified using PCR. (left) Marker, (middle) native λ DNA, (right) a plaque of λ-ARS317. (D) Purified λ-ARS317 DNA was detected by 0.6% agarose gel electrophoresis. (left) Marker, (middle) native λ DNA (right) purified λ-ARS317. Please click here to view a larger version of this figure.
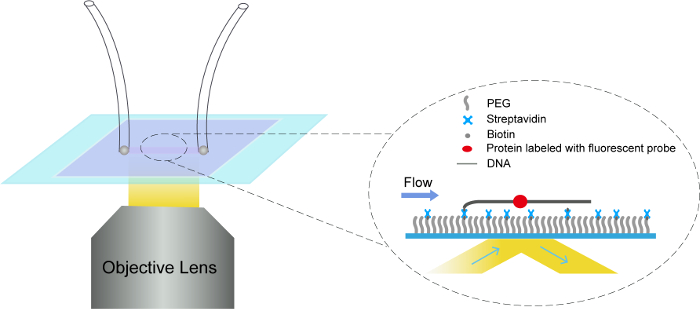
Figure 2: Schematic overview of objective-type TIRF and flow cell. Single molecule imaging assays were carried out based on the TIRFM combining with flow cell system. Our TIRFM was performed on an inverted microscope fitted with a 60X oil objective (numerical aperture = 1.49). DNA (gray line) was tethered on the coverslip in the flow cell through the biotin-streptavidin linkage, and stretched by the flow from left to right. A protein binding on the tethered DNA was indicated by a red-oval dot. Please click here to view a larger version of this figure.
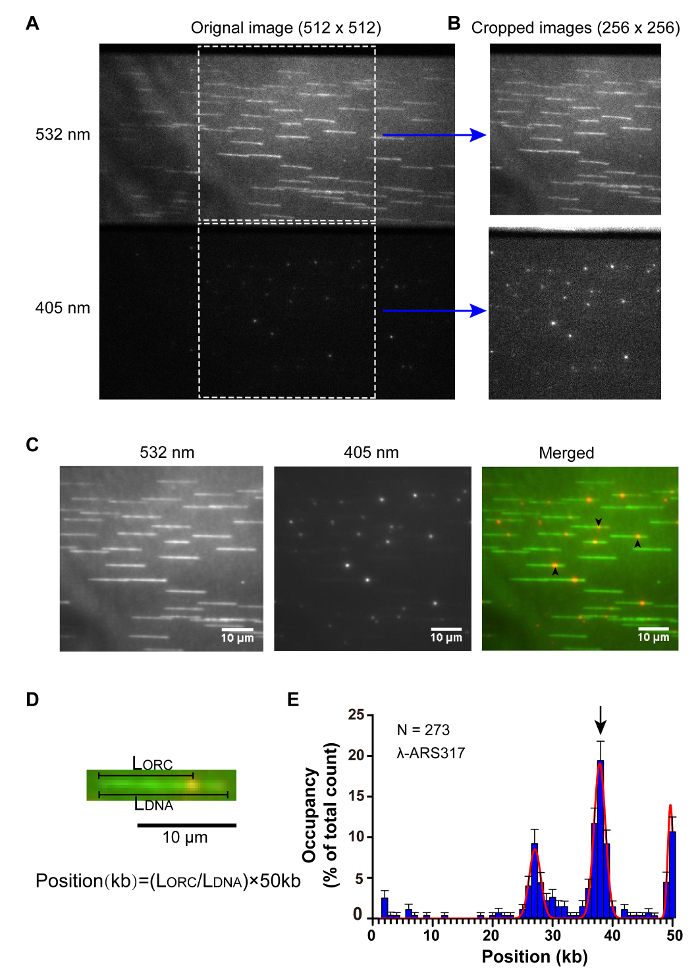
Figure 3: Qdot705-labeled ORC (1:1 molar ratio) binding on λ-ARS317. (A) ORC and λ-ARS317 DNA were observed simultaneously. (top) SYTOX Orange stained DNA was excited using a 532 nm laser and observed using the 550-613 nm transmission band of the quad-band band-pass filter. (bottom) Qdot705-labeled ORC was excited using a 405 nm laser and observed using the 663-743 nm transmission band of the quad-band band-pass filter. (B) Two 256×256 sub-stacks were cropped from the original 512×512 stack. (C) The images acquired using 61 sequential frames in the corresponding stacks. (left) SYTOX Orange stained DNA, (middle) Qdot705-labeled ORC, (right) merged image; three Qdot705-labeled ORC binding at ARS317 site on λ-ARS317 DNA substrates were marked using black arrows. (D) Illustration of the calculation of ORC binding position on DNA. LDNA means the DNA length, LORC means the distance from ORC binding site on DNA to the tethered end of DNA. (E) Histogram of ORC binding distribution on λ-ARS317 DNA. Gaussian fit to the data was indicated using the red solid line. Error bars indicate a 95% confidence interval based on 1000 bootstrap samples. N means the number of ORC molecules. ARS317 inserted site was indicated using a black arrow. Please click here to view a larger version of this figure.

Figure 4: Qdot-labeled ORC binding (3:1 molar ratio) on λ-ARS317. ORC and λ-ARS317 DNA were observed in the same way as in Figure 3A. (left) SYTOX Orange stained DNA was excited using a 532 nm laser. (middle) Qdot-labeled ORC was excited using a 405 nm laser. (right) merged images. Two Qdot-labeled ORC binding at ARS317 site on λ-ARS317 DNA substrates were marked using black arrows. Please click here to view a larger version of this figure.
