Nanobar design is recommended for probing positive curvature sensing proteins, which contains a half circle at each end with curvature defined by the nanobar width and one flat/zero curvature control locally at the center (Figure 2A,B). Successful formation of the SLB on nanobars results in evenly distributed lipid marker signals across the entire nanobar surface as shown in Figure 2C. Signals from multiple nanobars can be combined by averaging the individual nanobar images (Figure 2D) so that random variations between different nanobars can be minimized. An earlier electron microscopy study showed that cell plasma membrane formed conformal coating around the entire nanostructures24. Therefore, the synthetic lipid bilayer is believed to attach tightly on the nanostructure surface as well, which gives diffraction-limited lines on the 200 nm nanobar as observed in Figure 4E. In addition, the membrane fluidity on the nanobar-curved SLB can also be characterized by FRAP assay, where the fluorescent signal on a single nanobar can be bleached individually for membrane fluidity characterization (Figure 2E,F).
The formation of the SLB on nanobars requires the generation of SUVs similarly to the SLB formation on flat surfaces as reported earlier17. The lipid composition of SUVs determines the surface chemistry of the nanobar-curved SLB that can be altered for membrane labeling and different protein binding mechanisms. For example, Texas Red DHPE can be added in ~0.5 mol% in SUVs so that the enrichment of the membrane surface area on nanobars can be easily imaged by fluorescence microscopy (Figure 2C). Besides visualization, lipid composition can also be changed to facilitate protein binding on nanobar-curved SLBs. For example, to facilitate the anchorage of His-tagged proteins, nickel-nitrilotriacetic acid (Ni-NTA) can be brought to the SLB on the nanobars by incorporating 1,2-dioleoyl-sn-glycero-3-[(N-(5-amino-1-carboxypentyl) iminodiacetic acid) succinyl] (nickel salt) (18:1 DGS-NTA(Ni)) into SUVs (~1 mol%)25. Without this functional group on lipids, green fluorescence protein (GFP) cannot bind to the SLB with negatively charged PS, resulting in a darker bar-shaped hole at each nanobar location due to volumetric depletion of GFP signals in the solution (Figure 2G). In comparison, by adding 1 mol% 18:1 DGS-NTA(Ni) in the SLB, the His-tag labeled GFP can strongly anchor on the membrane to clearly follow the SLB shape curved by nanobars (Figure 2H). The lipid diffusion did not show a detectable difference upon protein binding as probed by FRAP assay (Supplementary Figure 1). It is worth noting that the FRAP measurement is based only on the intensity of the 0.5 mol% Texas Red DHPE in the SLB, which may not represent the 1 mol% DGS-NTA(Ni) used for the binding of the His-tag protein. In addition, charged lipids such as PS can enhance protein binding through electrostatic interaction26, while signaling lipids such as phosphatidylinositol (4,5)-bisphosphate (PIP2) can also be integrated in SUVs to facilitate specific protein binding (e.g., FCHo2, for which PIP2 is found essential for its membrane curvature sensing27).
The quality of SLB formation in terms of surface uniformity and membrane fluidity is critical for probing the protein curvature sensing ability on nanobars. There are three typical issues that may compromise SLB uniformity and fluidity. One is the binding of excess SUVs on the SLB due to inefficient washing that generates puncta both on the nanobars and on the flat membrane between the nanobars (Figure 3A,B). It significantly affects the quantification of protein binding on curved or flat surfaces on nanobars for curvature sensing assessment. Another issue is the unexpected introduction of air bubbles inside the PDMS chamber during the washing step, which may destroy the SLB along the trajectory of the air-liquid interface and leave scratch-like features easily identifiable with extremely low lipid signals on the nanobar surface (Figure 3C,D). Moreover, improperly cleaned nanobar substrates leave randomly distributed chemical residues on the surface that prevent lipid adsorption for SLB formation. It leads to the generation of membrane defects that may or may not be above the optical resolution for microscopy visualization (Figure 3E). However, membrane fluidity will be sensitively affected by membrane defect formation28 so that FRAP assay can be used to verify the surface cleanness (Figure 3F,G).
To demonstrate the characterization of curvature sensing protein using nanobar array, the typical F-BAR domain is compared with a recently identified intrinsically disordered region in FBP17 (IDRFBP17). F-BAR is fluorescently labeled with Alexa Fluor 488 tetrafluorophenyl ester dyes while IDRFBP17 is labeled with Alexa Fluor 647 C2 Maleimide. As shown in Figure 4A, both protein domains show increased accumulation on SLB-coated individual nanobars with 300 nm width. Here, the SLB contains 10% PS to electrostatically enhance the protein binding. The protein's curvature sensing ability can be identified by the preferential enrichment at the curved nanobar ends with higher fluorescent intensity than the signals at the flat nanobar centers, which is more obviously shown after image averaging from more than 100 nanobars (Figure 4B). The curvature sensing ability can be quantitatively reflected by calculating the end-to-center ratio on each nanobar (i.e., the fluorescence intensity at the nanobar end versus the nanobar center). As shown in Figure 4C, both proteins have a higher end-to-center ratio than lipids, which is around 1. When comparing the IDRFBP17 and F-BAR, it can be seen that the higher end-to-center ratio of IDRFBP17 (1.385 ± 0.011, mean ± SEM) indicates stronger curvature sensing than F-BAR (1.144 ± 0.004, mean ± SEM).
To examine the range of curvatures that can be sensed by the proteins, the SLB is generated on nanobar arrays with gradient widths from 200 nm to 600 nm (Figure 4D) where lipids showed homogeneous coating on different-sized nanobars (Figure 4E). Three proteins (IDRFBP17, F-BAR, and F-BAR+IDRFBP17) are incubated on the gradient nanobar array, and all of them showed preferential accumulation at the nanobar-end curved membrane sites when the curvature decreased below 400 nm in diameter (Figure 4E,F). Among these three protein domains, IDRFBP17 gives the highest nanobar-end density, indicating its strongest curvature sensing ability, while F-BAR shows the lowest value (Figure 4F). Furthermore, the binding curve of the curvature sensing protein can also be plotted by gradually increasing the protein concentration (Figure 4G), which shows that IDRFBP17 has a strong cooperative curvature sensing ability. When fitting its binding curves to the Hill equation, the KD is found in the µM range (0.6 ± 0.1 µM) and the Hill coefficient (H) is between 2 and 3, suggesting an ultrasensitive binding of IDRFBP17 to nanobar-induced membrane curvatures. The sharper the curvature, the higher the binding capacity at equilibrium.
The dynamic membrane diffusion and membrane association of curvature sensing proteins around the nanobar-shaped SLB sites can also be studied by FRAP. Compared to the fast recovery of lipid signals on nanobars (Figure 5A,E), the F-BAR could not recover within 2 min (Figure 5B,E), suggesting its significantly decreased membrane mobility and association dynamic at the curved membrane sites. Surprisingly, different from the behavior of F-BAR, IDRFBP17 signal showed obvious recovery within the same time frame (Figure 5C,E), indicating the dynamic nature of IDRFBP17 accumulated at the curved membranes. However, after washing away the unbound IDRFBP17 in the solution, the same recovery in the IDRFBP17 channel couldn't be observed as before (Figure 5D,E). It indicates that the recovery is dominated by the exchange with the solution-containing IDRFBP17 molecules and less by lateral diffusion of the membrane-bound molecules.
Overall, these results demonstrate that this nanobar-SLB system is a powerful tool to characterize membrane curvature-sensing proteins in vitro.
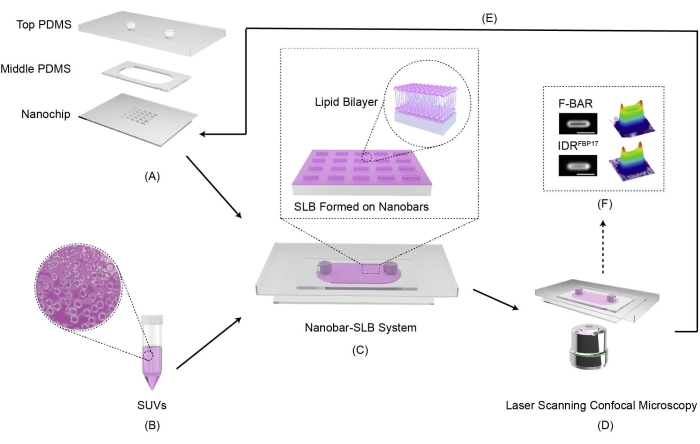
Figure 1: Illustration of the nanobar-SLB system. (A) The PDMS chamber consists of the top and middle layers with the cleaned nanochip assembled on it. (B) The Texas Red DHPE labeled SUVs with zoomed-in detail. (C) The intact model of the nanobar-SLB system and the zoom-in area illustrates that the SLB forms on nanobars with a uniform single layer of the lipid bilayer. (D) Imaging and characterization process under laser scanning confocal microscopy. (E) Cleaning the nanochip for further reuse. (F) Imaging processing for protein curvature sensing quantification. Please click here to view a larger version of this figure.
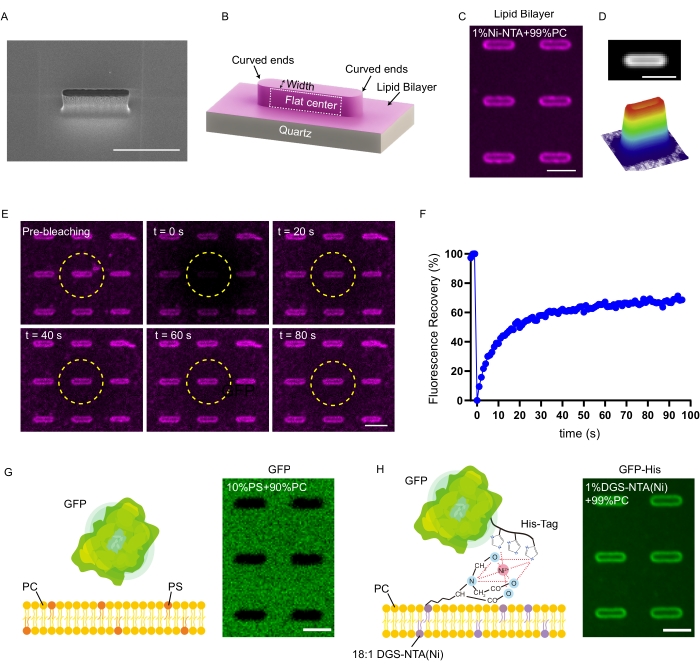
Figure 2: The formation of the curved SLB on nanobars with protein binding. (A) SEM image of an individual nanobar. (B) Schematic depiction of the nanobar-SLB system. (C) Fluorescent images of the SLB labeled with Texas Red DHPE on the nanochip. (D) Average image of SLB distribution on nanobars (top) and a 3D surface plot of the average image (bottom). This figure has been modified from Su et al.22 and reproduced with permission. (E) Time-lapse imaging of the SLB FRAP on the nanobar. The bleaching region is indicated by a yellow dashed circle. (F) Normalized nanobar intensity plot by FRAP measurement. (G) Schematic depiction of GFP cannot bind to the SLB with 10 mol% negative charged PS (left) and a corresponding representative image on the right. (H) Schematic depiction of His-tag labeled GFP binded to the SLB with 1 mol% DGS-NTA(Ni) (left) and a corresponding representative image on the right. Scale bars: 2 µm. Please click here to view a larger version of this figure.
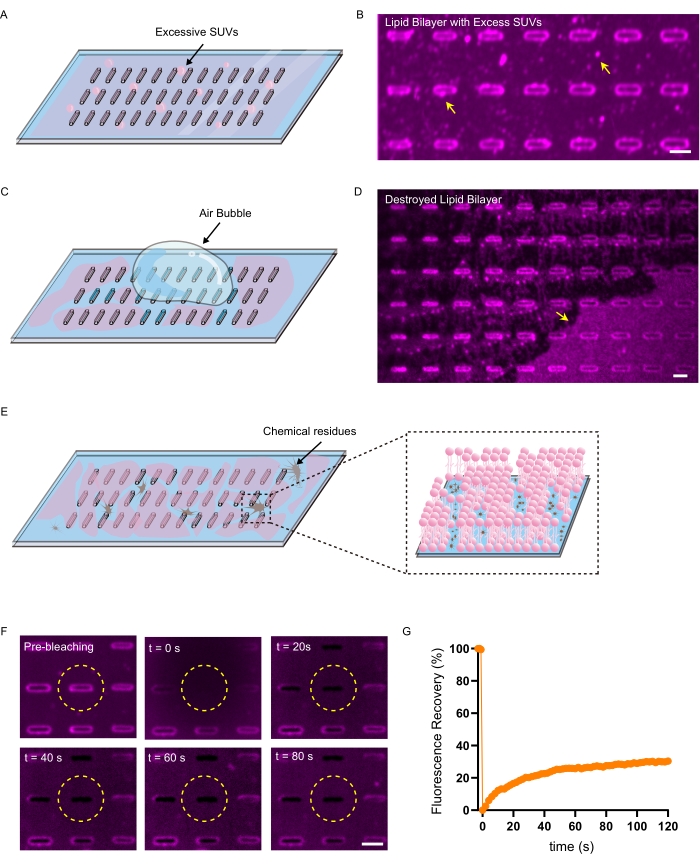
Figure 3: Typical issues that compromise the SLB quality. (A) Schematic of SLB preparation without efficient washing. (B) Representative image of the SLB bind with excess SUVs. The yellow arrows show the puncta which are excess SUVs on the SLB surface. (C) Schematic of SLB preparation with air bubbles injected. (D) Representative image of SLB destroyed by the trajectory of the air-liquid interface. The yellow arrow shows the incomplete SLB on the substrate. (E) Schematic of SLB preparation with uncleaned nanobar substrates. The zoom-in area shows a discontinuous lipid bilayer due to the surface residues prevent lipid adsorption. (F) Time-lapse imaging of SLB FRAP analysis on uncleaned nanobar substrates. The bleaching region is indicated by a yellow dashed circle. (G) Normalized nanobar intensity plot by FRAP measurement. Scale bars: 2 µm. Please click here to view a larger version of this figure.
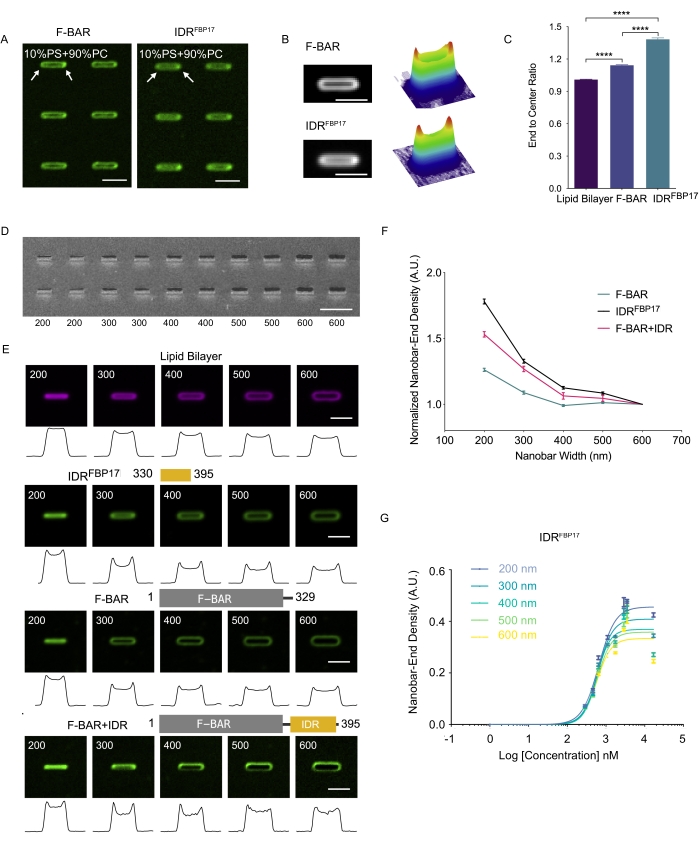
Figure 4: Protein curvature sensing characterization by the nanobar-SLB system. (A) Fluorescent images of F-BAR (left) and IDRFBP17 (right) accumulate on SLB-coated nanobars (10% PS and 90% PC). Scale bar: 2 µm. (B) Average images of F-BAR (left top) and IDRFBP17 (left bottom) accumulate on nanobars and corresponding 3D surface plots (right top and right bottom, respectively). Scale bar: 2 µm. (C) End-to-center ratio of the lipid bilayer, F-BAR and IDRFBP17 fluorescence intensity around the 300 nm width nanobar. A Welch's t-test was performed for statistical analysis. ****p < 0.0001. (n = 3) (D) SEM image of gradient nanobar arrays with widths from 200 nm to 600 nm. Scale bar: 5 µm. (E) Annotation of protein construct (top), average images of lipid bilayer, IDRFBP17, F-BAR, and F-BAR+IDRFBP17 distribution on gradient nanobars with width ranging from 200 nm to 600 nm (middle) and intensity profiles along the nanobars of each average image (bottom). Scale bar: 2 µm. (F) IDRFBP17, F-BAR, and F-BAR+IDRFBP17 normalized nanobar-end density quantified with different widths of nanobars respectively. Each data is shown as the mean ± SEM (n ≥ 60). (G) The binding curve of IDRFBP17 fitted with the Hill equation. Each data is shown as the mean ± SEM (n = 3). This figure has been modified from Su et al.22 and reproduced with permission. Please click here to view a larger version of this figure.
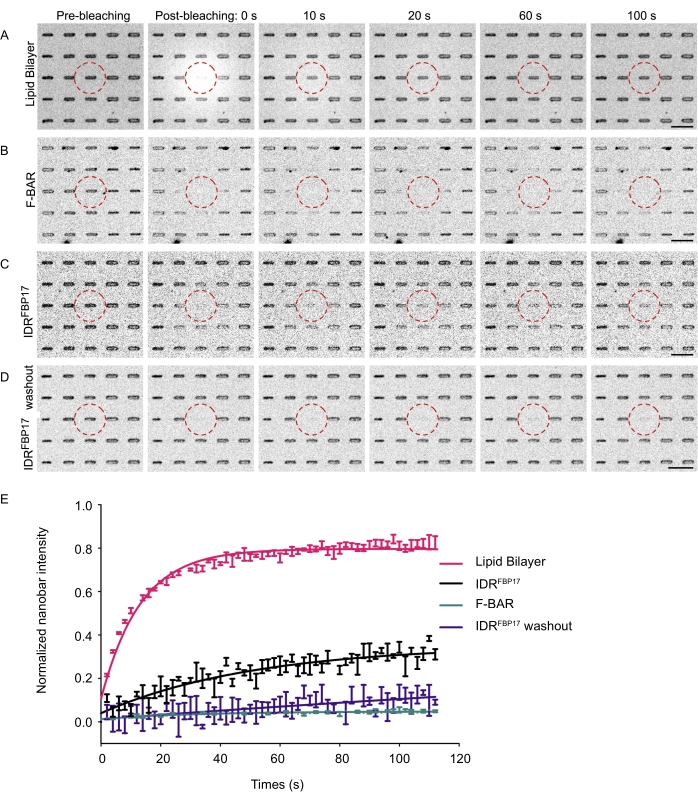
Figure 5: The membrane-binding dynamics of curvature sensing proteins around the nanobar. (A–D) Time-lapse imaging of the lipid bilayer (A), F-BAR (B), IDRFBP17 (C), and IDRFBP17 washed out (D) FRAP analysis on the nanobar. The bleaching region is indicated by a red dashed circle. Scale bar: 5 µm. (E) Normalized nanobar intensity plot by FRAP measurement. This figure has been modified from Su et al.22 and reproduced with permission. Please click here to view a larger version of this figure.
Supplementary Figure 1: Membrane diffusion before and after adding protein. (A) Time-lapse imaging of FRAP test on the POPC lipid bilayer with 1 mol% 18:1 DGS-NTA(Ni) and 0.5 mol% Texas Red DHPE around the nanobar before (top) and after (bottom) adding GFP-His protein. The bleaching region is indicated by a yellow dashed circle. (B) Normalized nanobar intensity plot by FRAP measurement. Scale bars: 2 µm. Please click here to download this File.
