Using the protocol described above, we detected the sites of interaction of two ER-anchored lipid transfer proteins, ORP5 and ORP8, and assessed their occurrence at ER membrane subdomains in contact with other organelles, in particular, with the mitochondria. For that, the mitochondrial network in HeLa cells was stained with a red mitochondrial marker, and ORP5-ORP8 PLA green spots were detected after fixation using the primary antibodies anti-ORP5 and anti-ORP8, whose specificity was previously tested by immunofluorescence15. Confocal images showed that endogenous ORP5-ORP8 PLA interactions in the HeLa cells occurred in the reticular ER, cortical ER, and in ER subdomains in close contact with mitochondria, commonly referred to as mitochondria-associated ER membranes (MAMs; Figure 2A). In order to quantify the ORP5-ORP8 PLA interactions occurring at MAMs, the distance of each PLA spot from mitochondria was evaluated using 3D image analysis. Using a distance threshold of 380 nm, 3D imaging analysis revealed that about 50% of endogenous ORP5-ORP8 PLA interactions were detected at MAMs (Figure 2A,B). The other 50% of interactions were distributed between the cortical and reticular ER15. To validate the specificity of the PLA, ORP5-ORP8 PLA experiments were performed either in cells treated with ORP5-targeting and ORP8-targeting siRNAs (negative control) or in cells co-overexpressing ORP5 and ORP8 (positive control). ORP5 and ORP8 downregulation induced a massive decrease in the total number of PLA signals (at MAMs, at the reticular ER, and at the cortical ER; Figure 2A,C), while their co-overexpression resulted in an increase in PLA (Figure 2A,C), confirming the specificity of the ORP5-ORP8 PLA. However, interestingly, the percentage of ORP5-ORP8 PLA interactions occurring at MAMs in cells co-overexpressing ORP5 and ORP8 was similar to that observed in cells where these proteins were expressed at endogenous levels (Figure 2B), supporting their existence as a complex at MAMs. To further confirm the presence of ORP5 and ORP8 at ER membrane subdomains in close contact with mitochondria, additional quantitative PLA analyses between ORP5 or ORP8 and the outer mitochondrial membrane protein PTPIP51, a known binding partner of ORP5 and ORP813, were carried out. PLA signals were detected in both ORP5-PTPIP51 and ORP8-PTPIP51 couples, and their average numbers were similar to the ORP5-ORP8 PLA couple, confirming ORP5 and ORP8 localization at ER-mitochondria MCSs (Figure 3).
Finally, in recent works from our lab, by using a similar approach in HeLa cells transfected with PHPLCd-RFP or mcherry-PLN1 to mark the PM or LD, respectively, we were able to analyze the occurrence of ORP5-ORP8 PLA interactions at ER-PM contacts and to identify a three-way contact site between mitochondria, the ER, and LDs (Figure 4)15,17.
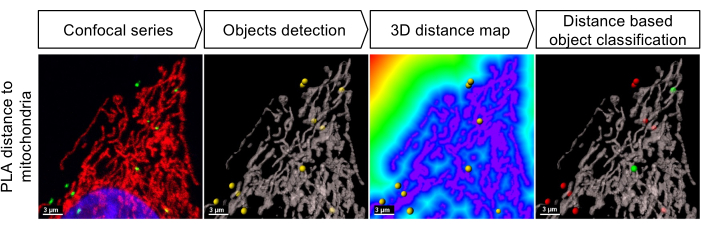
Figure 1: Workflow for the identification of PLA signals in close proximity to the mitochondria. First, the confocal stacks are segmented to identify the PLA foci (spots) and the mitochondrial network (surfaces). Then, 3D distance maps are computed toward the outside of the surfaces (mitochondria), allowing the measurement of the distance of each spot from the closest mitochondrion. Finally, PLA spots are classified into two populations (red and green) based on a proximity threshold of 380 nm established based on the precision of the detection system. This figure has been modified fromMonteiro-Cardoso et al.15. Please click here to view a larger version of this figure.
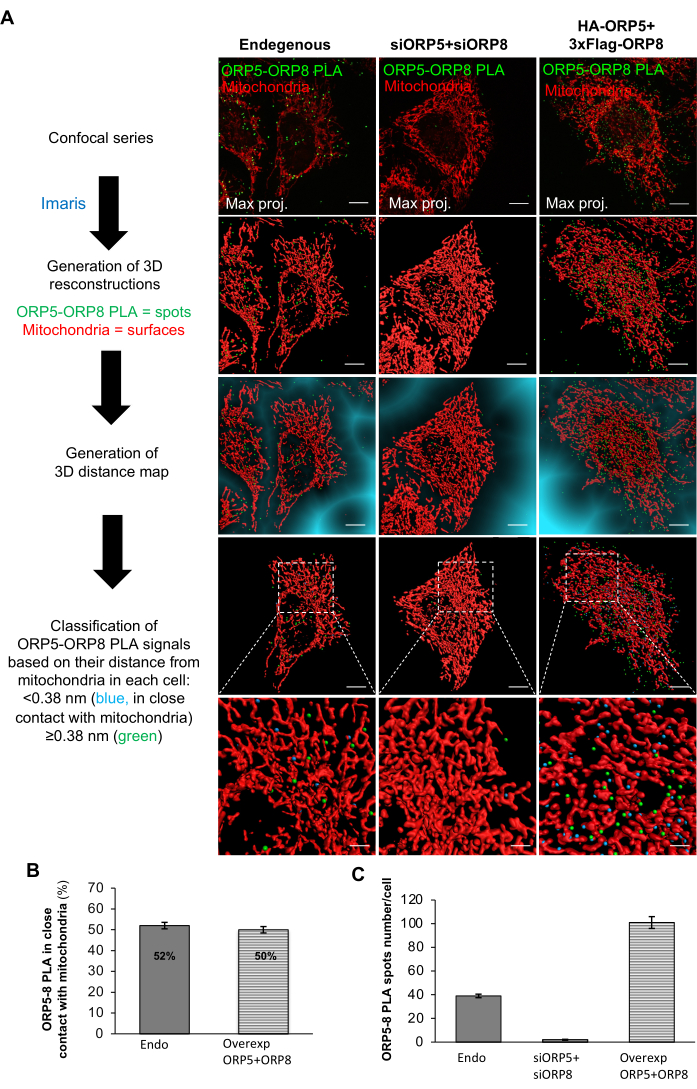
Figure 2: Representative images of the ORP5-ORP8 complex expressed at endogenous levels localizing at ER-mitochondria contacts. (A) ORP5-ORP8 PLA confocal series and respective 3D reconstitutions. Scale bars: 10 µm and 2 µm (expanded images). (B) Quantification of ORP5-ORP8 PLA foci in close contact with mitochondria. Endo, n = 33 cells, and Overexp ORP5+ORP8, n = 27 cells. Data are presented as mean ± SEM (standard error of the mean). This image has been modified from Monteiro-Cardoso et al.15. (C) Quantification of total ORP5-ORP8 PLA foci. Endo, n = 38 cells, siORP5+siORP8, n = 38 cells, and Overexp ORP5+ORP8, n = 35 cells. Data are presented as mean ± SEM (standard error of the mean). This image has been modified from Monteiro-Cardoso et al.15. Please click here to view a larger version of this figure.
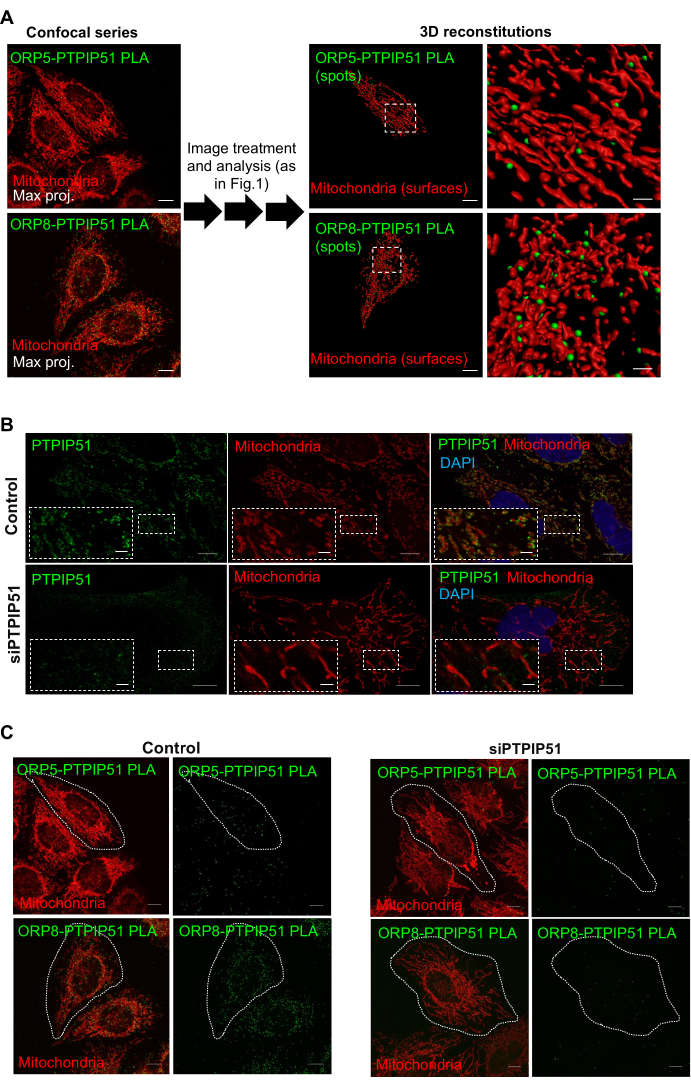
Figure 3: Representative images of PLA signals between ORP5 or ORP8 with the mitochondrial protein PTPIP51. (A) Confocal series were used to generate 3D reconstitutions and distance maps to confirm that ORPR5 or ORP8 PLA interactions with PTPIP51 occur at ER-mitochondria close contacts. Scale bars: 10 µm and 2 µm (expanded images). (B) PTPIP51 immunofluorescence confocal images (single focal plane) in control and HeLa cells treated with siRNA targeting PTPIP51. Scale bars: 10 µm and 2 µm (expanded images). (C) ORP5-PTPIP51 and ORP8-PTPIP51 PLA confocal images in control and HeLa cells treated with siRNA targeting PTPIP51. Scale bar: 10 µm. Please click here to view a larger version of this figure.
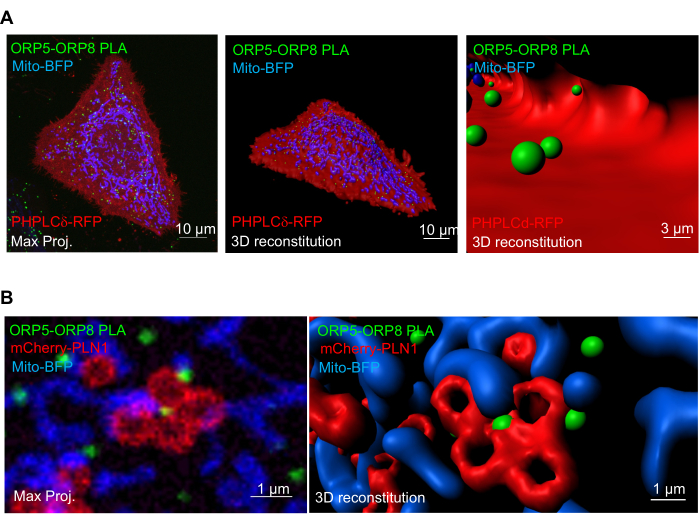
Figure 4: Examples of 3D reconstructions used to localize the ORP5-ORP8 PLA complex at ER-PM and ER-mitochondria-LD contacts. (A) Confocal image and respective 3D reconstitution of a HeLa cell with ORP5-ORP8 PLA, represented by the green spots, the PM marked with PHPLCd-RFP, represented by the red cell, and mitochondria marked with Mito-BFP, represented by blue surfaces. Left and center images: representation of the whole HeLa cell. Right image: representation of a zoomed region inside the cell. (B) Confocal image and respective 3D reconstitution of a zoomed-in region of a HeLa cell with ORP5-ORP8 PLA, represented by the green spots, LDs marked with mCherry-PLN1, represented by the red surfaces, and Mito-BFP, represented by the blue surfaces. Please click here to view a larger version of this figure.
