The 3D T/L organoid model was previously established and demonstrated here by implementing commercially purchased NHDF (n=3, 3 organoids per donor, NHDF were used at passages 5-8). The model workflow is summarized in Figure 1. Figure 2 shows representative phase-contrast images of NHDF culture during the pre-expansion in T-75 flasks (Figure 2A) as well as at the beginning and after 5 days of culture in the 2D expansion step in 10 cm cell culture dishes (Figure 2B). Figure 2C (representative phase-contrast images) demonstrates the confluence of NHDFs during 2D stimulation after 14 days of culture in the 10 cm cell culture dishes. Subsequently, the cell sheets were manually detached and simultaneously rolled into 3D rod-like organoids (at day 0, organoid length is approximately 6 cm, width approximately 0.4 cm), which were cultivated under 10 % static tension for 14 days (Figure 3, day 0 and day 14, representative macroscopic images). By day 14 of the 3D maturation process, the organoids displayed a glistening white appearance, contracted, and appeared denser than the initial organoids. The wet weight of the organoids was measured at the time of formation (day 0) and again on day 14 of the 3D maturation step (Figure 4). A significant reduction in the wet weight was observed, indicating contraction and ECM structural reorganization within the organoids during the time span of this protocol step. In addition to weight change, due to the contraction, the organoid length also reduced (at day 14, the organoid length is approximately 3.5 cm, and the width is approximately 0.3 cm). In this process, the metal pins loosened up and had to be re-fixed; hence, during the 14 days of 3D maturation, we recommend monitoring the organoids frequently. To evaluate the morphology of the organoids, longitudinal sections were obtained from NHDF organoids collected on day 1 and day 14 of the 3D maturation step and subjected to H&E staining (Figure 5). On day 1, the organoids displayed disconnected layers, primarily composed of cells surrounded by low amounts of ECM. Furthermore, the cells within the organoid layers were not aligned along the longitudinal organoid axis used as direction for the applied static tensile load. The NHDF nuclei exhibited a round shape with varying sizes and shapes. H&E analysis of day 14 organoids revealed a noticeable increase in the eosin signal, indicating ECM deposition during the 3D maturation process. At this time point, the organoids exhibited fused layers, with areas containing aligned cells. Cells were frequently in groups; however, they were also organized in cell rows in some locations. The majority of the nuclei had similar sizes and occasionally were elongated.
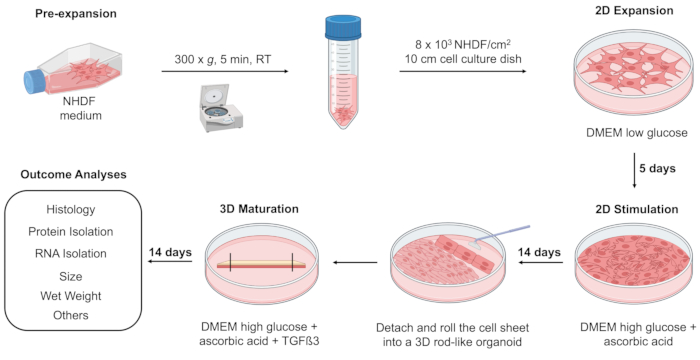
Figure 1: Cartoon of 3D T/L organoid model composed of 3 steps: 2D expansion, 2D stimulation, and 3D maturation. Initially, NHDFs were pre-expanded in a T-75 flask until they reached 70%-80% confluence. Subsequently, NHDFs were cultivated using Dulbecco's Modified Eagles Medium (DMEM) low glucose medium in a 10 cm cell culture dish for 5 days (2D expansion). The cells were then stimulated with DMEM high glucose medium supplemented with ascorbic acid for 14 days of culture (2D stimulation). Following, NHDF were detached from the 10 cm adherent dish, rolled into a 3D rod-like organoid, transferred, and fixed in a 10 cm non-adherent cell culture dish filled with DMEM high glucose medium supplemented with ascorbic acid and Transforming Growth Factor beta 3 (TGFß3) for 14 days (3D maturation). The formed 3D organoids may undergo an assessment of wet weight, histological evaluation, RNA and protein isolation, and other analyses (e.g., transmission electron microscopy, biomechanical measurements) at different time points such as days 0, 1, and 14. The cartoon was generated in BioRender software. Please click here to view a larger version of this figure.
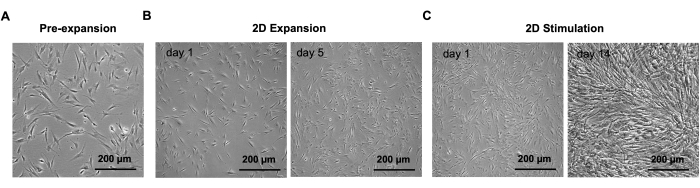
Figure 2: Representative phase-contrast images of NHDFs during pre-expansion, 2D expansion, and 2D stimulation steps. (A) NHDFs during pre-expansion in T-75 flask. (B) NHDFs at days 1 and 5 of the 2D expansion step in a 10 cm cell culture dish. (C) NHDFs at days 1 and 14 of 2D stimulation step in a 10 cm cell culture dish. Scale bars: 200 µm. Images were taken with an inverted microscope equipped with a high-resolution camera. Please click here to view a larger version of this figure.

Figure 3: Gross morphology of NHDF 3D organoids. Representative macroscopic images of NHDF organoids at days 0 and 14 of the 3D maturation step. Scale bar: 1 cm. Images were taken with a mobile phone camera. Please click here to view a larger version of this figure.
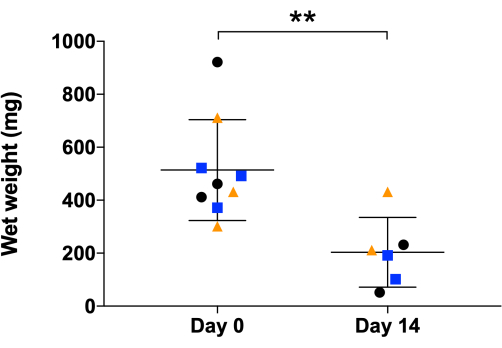
Figure 4: Wet weight analysis of NHDF 3D organoids. Organoids' wet weight measurement was at day 0 and day 14, the end of the 3D maturation process. The graph shows mean values and standard deviations (NHDF n = 3, 3 organoids/donor at day 0 and 2 organoids/donor at day 14 due to 1 organoid/donor taken for histology at day 1), each dot represents individual organoids while each colored shape represents a different individual donor. Data is presented as mean ± standard deviation, and an unpaired parametric t-test was used. Statistical differences: **p ≤0.01. Please click here to view a larger version of this figure.
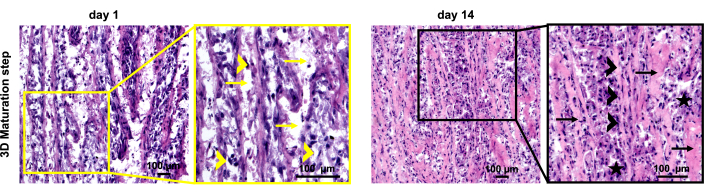
Figure 5: Histological analysis of NHDF 3D organoids. Representative images of hematoxylin and eosin (H&E) stained sections of NHDF organoids during the 3D maturation process at days 1 and 14. Right image – low magnification images, left image – magnified view with indicators as follows: yellow arrows – disconnected layers; yellow arrowheads – nuclei with varying sizes and shapes; black arrows – fused layers and ECM deposition; black arrowheads – cell rows, nuclei with similar size and elongation; black stars -cells in groups. Scale bars: 100 µm. Please click here to view a larger version of this figure.
