מספר המבנים גבישיים של חלבונים וקומפלקסי חלבונים גדל בקצב מהיר בשנים האחרונות. הם מציגים תמונות לא יסולא בפז של הארגון המבני של חלבונים אלה ולספק בסיס לניתוח תפקוד המבנה. עם זאת, הדינמיקה של חלבונים ושינויי קונפורמציה, שהם חיוניים לתפקודם, מתגלה לעתים רחוקות על ידי קריסטלוגרפיה באמצעות קרן רנטגן. Cryo-electronmicroscopy, לעומת זאת, הוא מסוגל ללכוד מתחמי חלבון וחלבון בתצורות שונות, אך בדרך כלל לא יכול לפתור את שינויי קונפורמציה לרמת מבנה משני 1. דינמיקה קונפורמציה של חלבונים בתמיסה בפרטים אטומיים יכולה להיפתר רק על ידי תמ"ג, אך בשיטה זו היא עדיין מוגבלת לחלבונים בגדלים קטנים יחסית (בדרך כלל ≤ 30 KDA) וזקוק לריכוז גבוה של חלבונים (≥ 100 מיקרומטר), דבר המקשה ניסויים עם חלבונים נוטים oligomerization או צבירה 2. שיטה אחת שהוא מסוגל לגשר בין קריסטלוגרפיה ברזולוציה הגבוהה של קרני רנטגן, cryo-electronmicroscopy ושאינו מוגבל על ידי גודל חלבון או ריכוז הוא מימן-1 אמיד H / 2 H-חליפין (HX) בשילוב עם ספקטרומטריית מסה (MS). בשנים האחרונות שיטה זו התפתחה לכלי אנליטי חשוב לניתוח דינמיקת חלבון, קיפול חלבון, חלבון יציבות ושינויי קונפורמציה 3-5. הבסיס המולקולרי של שיטה זו הוא באופי יציב של עמוד השדרה מימנים אמיד בחלבונים, שיחליפו עם אטומי דאוטריום כאשר החלבון ממוקם בD 2 O פתרון. העלייה שלאחר מכן במסת חלבון לאורך הזמן נמדדת עם רזולוציה גבוהה טרשת נפוצה.
בפפטידים בלתי מובנים קצרים HX רק תלוי בטמפרטורה, ריכוז זרז (OH -, H 3 O + כלומר pH, ראה איור 3) ורשתות צד חומצת אמינו של שיירים סמוכים בשל אינדוקטיביים, חתולהשפעות alytic וסטריות. השפעות אלה על ch k שער חליפין כימיים הפנימי היו לכמת באלגנטיות על ידי באי et al. 6 ותכנית זמינה (באדיבות ז''אנג), המחשבת ch k לכל חומצת אמינו בתוך פוליפפטיד התלוי בpH וטמפרטורה. ב-pH הניטרלי וטמפרטורות הסביבה ch k הוא בסדר הגודל של 10 1 -10 3 שניות -1. בחלבונים מקופלים HX יכול להיות 2-9 סדר גודל איטי בעיקר בשל קשרי מימן במבנה משני ובמידה קלה בשל גישה מוגבלת של התייבשות OH – יונים לפנים של חלבון מקופל ומהודק. HX בחלבוני ילידים לכן מסבך, החלפה חלקית או גלובלי התגלגלות כימית וקפל שוב למצב הטבעי על פי משוואה (1) ושעיר חליפין שנצפו k תצפיות תלויות באופ k שיעור פתיחה, CL k שער סגירה ואת חילופי הכימיים הפנימיים rate k ch על פי משוואה (2).
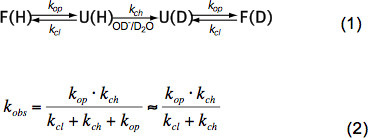
בתנאי מדינה האם של אופ k הוא הרבה יותר קטן מאשר ch k ויכול להיות מוזנח במכנה. ישנם שני משטרי חילופי קיצוניים בשם EX1 וEX2. אם CL k הוא הרבה יותר קטן מאשר ch k (EX1) השיעור שנצפה הוא כמעט שווה לשיעור הפתיחה וHX מאפשר תצפית מיידית של התגלגלות של אלמנט מבני. כגון משטר חליפין, שבו כל תמורת הפרוטונים אמיד מייד עם פתיחתו של היסוד המבני, היא לצפות בקלות בטרשת הנפוצה על ידי הפצת bimodal של פסגות איזוטופ 7. אם CL k הוא הרבה יותר גדול מch k (EX2) השיעור שנצפה הוא יחסי ch k לפיה המתמיד המידתיות שווה לאיזוני התגלגלות קיפולי אופ K u = k הקבוע </sub> / K CL. בתנאים אלה, אירועי פתיחה וסגירה רבים הם הכרחיים לפני כל החלפת הפרוטונים אמיד לdeuterons, מה שמובילים לעלייה הדרגתית במסה ממוצעת בעת החלוקה איזוטופי נשארה בערך אותו הדבר. משטר EX2 מאפשר קביעת האנרגיה החופשית של התגלגלות ΔG u ולכן היציבות של אלמנט מבני. בתנאי מדינה האם של משטר EX2 הוא נפוץ ביותר. עלייה של pH ובנוסף סוכני chaotropic יכולה להזיז את מנגנון החליפין לEX1. לכן, HX-MS יכול לשמש כדי לחקור התרמודינמית וכן פרמטרים הקינטית של קיפול חלבונים ושינויי קונפורמציה.
כפי שצוין לעיל HX הוא במהותו pH וטמפרטורה תלויה ואת חילופי זמן מחצית החיים של פרוטון נחשף ממס לחלוטין מהקבוצה אמיד עמוד השדרה הוא בין 5-400 אלפיות שני ב-pH הפיזיולוגי (pH 7.6) ו30 מעלות צלזיוס, אבל 10 דקות ל> שעות 15 עם ממוצע של> שעה 2 ב-pH 2.9 ו0 °C (פרט לפרוטון של הקשר אמיד עמוד השדרה הראשון של פוליפפטיד, שמחליף עם זמן מחצית חיים של CA. 1-2 דק '). תחת תנאים כאלה איטיים החלפה אפשר לעכל את המדגם באמצעות פרוטאזות (למשל פפסין) הפועלים בתנאים אלה, עם יציאה החוצה לאבד את כל המידע הכלול בdeuterons המשולב. מאז כניסתה של מערכת העיכול פפטי בתנאי החלפה איטיים, ניתן לנתח לא רק את קינטיקה HX הכוללת של חלבונים באורך מלא, אבל HX יכול להיות מקומית לאזורים הספציפיים 8,9. רזולוציה מרחבית כרגע מוגבלת לגודל של שברים פפטי שנוצרו, שהוא באופן כללי בין 10-30 שאריות. עם זאת, שברים חופפים שנוצרו בשל האופי ספציפי של מחשוף ידי פפסין יכולים להוביל לעלייה ברזולוציה מרחבית. בנוסף, כמה פרוטאזות אחרות נמצאו להיות פעילים בתנאים להרוות, לעומת זאת, הרבה פחות יעיל מאשר עכלן 10. increa נוספתניתן להגיע se של רזולוציה מרחבית על ידי פיצול של פפטידים בשלב הגז על ידי שיטות ששמרו את דפוס deuteration כגון ניתוק לכידת אלקטרונים (ECD), ניתוק אלקטרון העברה (ETD) ודיסוציאציה multiphoton אינפרא אדום (IRMPD) 11-13. טכניקות אלו למנוע האובדן של רזולוציה מרחבית בשל הגירת intramolecular פרוטון ("ערבול"), אשר נצפתה על ידי ניתוק הנגרם התנגשות (CID) את טכניקת הפיצול הנפוצה ביותר. עם זאת, שיטות אלו דורשות אופטימיזציה לכל פפטיד בודד ולכן היא עדיין מאתגרת למדי.
HX-MS כבר בשימוש כדי לנתח אינטראקציות החלבון ליגנד וחלבונים כולל הרכבה הקופסית נגיפית 14-17. חלבון התגלגלות וקיפול מחדש, כמו גם שינויי קונפורמציה מושרה טמפרטורה נחקרו 7,18,19. זירחון וקונפורמציה הקשורות למוטצית חומצה אמינית אחת משתנים 16,20 וnucleotשינויי ide-induced נותחו 21,22. לכן, שיטה זו נראית מתאימה באופן אידיאלי לניתוח הרכבה ודינמיקה של מכונות מולקולריות. מועמד אחד אטרקטיבי, מנגנון שהוא עניין כללי גדול, הוא מורכב מלווה Hsp90.
