We performed an assay to see if we could detect a colorimetric readout of the association of PXR and steroid receptor coactivator-1 (SRC-1). Since yeast has significant sterol production, it has previously been shown that lacZ expression in yeast can be induced without the need for additional exogenous ligand. We found that lacZ expression (blue colonies) is also induced in the yeast strain transformed with PXR and SRC-1; however, there is no induction of LacZ expression (white colonies) in yeast transformed with empty vectors, PXR or SRC-1 individually (Figure 3). To determine whether ketoconazole (25 μM) disrupted PXR and SRC-1 interaction in yeast, replica plates containing ketoconazole were soaked with nitrocellulose and X-gal filter, lift, and β-galactosidase liquid assays were performed. We show that ketoconazole disrupts PXR and SRC-1 interactions in yeast since all the colonies from the replica filter were now white (which was also shown by the significantly reduced β-galactosidase activity by liquid enzymatic assays) (Figure 4A). We had previously shown in mammalian assays that the PXR mutant (T248E/K277Q) can be activated by a strong ligand (e.g. rifampicin) but is immune to the antagonistic effects of ketoconazole17. Similarly, when we engineered the PXR double mutant (T248E/K277Q) in the yeast plasmid and then performed yeast transformations with SRC-1, we were able to show that the colonies exposed to ketoconazole still retain LacZ expression (Figure 4B).
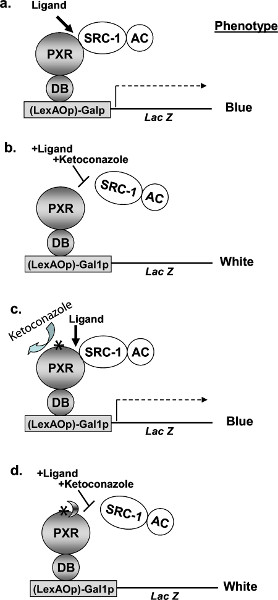
Figure 1. Schematic illustrating the principles of the yeast two-hybrid (Y2H) assay. The yeast assay has been established as a functional assay that allows for isolation of PXR mutants that are resistant to ketoconazole mediated inhibition of SRC-1 interaction. A, rifampicin-induced two-hybrid interaction of LexA-DB-hPXR fusion protein with SRC-1 fused to GAL4-activation domain (GAL4AC-SRC-1). The interaction results in blue colonies in an X-gal assay because of the LacZ reporter. B, presence of ketoconazole disrupts the rifampicin-induced interaction of hPXR with SRC-1. X-gal assay results in white colonies. C, presence of a mutation (indicated with an asterisk) in hPXR that renders it immune to the action of ketoconazole will yield a blue colony. D, a further modification of our assay in the future could incorporate additional second site suppressor mutations (illustrated with half moon) nullifying the effect of first mutation that could result in the disruption of hPXR-SRC-1 interaction and hence a white colony in the X-gal assay. LexAOp, LexA operon; Galp, Gal4 promoter, Lac Z, beta-glalactosidase; DB, LexAOp DNA-binding domain of Galp; AC, activation domain; *, mutation(s). (Reproduced from reference6). Click here to view larger figure.
Figure 2. Experimental Flow Chart.
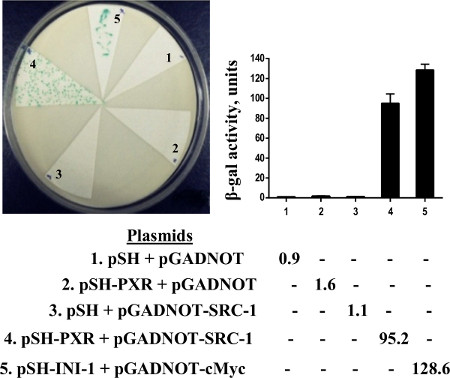
Figure 3. X-gal, lift assay, and β-gal liquid assay. Yeast was transformed with indicated plasmids and plated colonies subjected to X-gal lift assay (left panel) and β-gal liquid assay (right panel). pSH empty vector + pGADNOT empty vector (lane 1); pSH-PXR + pGADNOT empty vector (lane 2); pSH empty vector + pGADNOT-SRC-1 (lane 3); pSH-PXR + pGADNOT-SRC-1 (lane 4); 5. pSH-INI-1 + pGADNOT-c-Myc (positive control; lane 5). (reproduced from reference6).
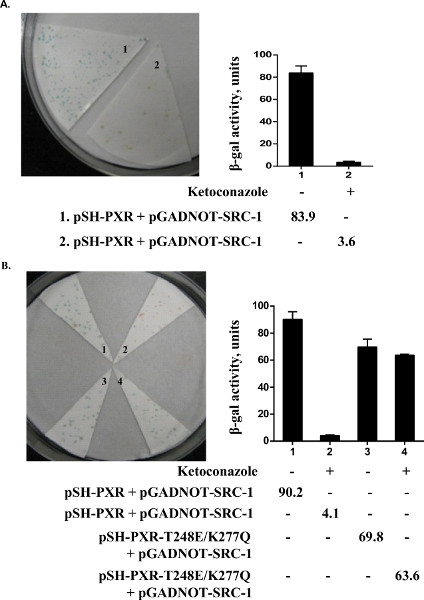
Figure 4. Ketoconazole disrupts wild-type but not mutant PXR association with coactivator, SRC-1. Yeast colonies were replica in plates containing vehicle (0.2% DMSO; lane 1) or ketoconazole (25 μM; lane 2). X-gal lift assay (left panel) and β-gal liquid assay (right panel) were then performed. Wild type PXR (A, B line 1 and 2) and ketoconazole mute mutant T248E/K277Q (B line 3 and 4) were shown in this figure. (reproduced from reference6).
