Dit artikel geeft een betrouwbare test om de relatieve expressie van celoppervlak membraaneiwitten zoals ionenkanalen tot expressie gebracht in recombinante cellen met de aanwezige flowcytometrie techniek melden. Ionenkanalen porie-vormende membraaneiwitten die verantwoordelijk zijn voor het regelen van elektrische signalen door het poorten van de stroming van ionen door de celmembraan. Zij worden ingedeeld door de activatie mechanisme, natuur, en de selectiviteit van de ion soorten op doorreis in de poriën waar ze worden gelokaliseerd. Op cellulair en weefsel niveaus, de macroscopische ion fluxen door middel van ion kanalen zijn het product van biofysische (gating en permeatie), biochemische (fosforylering), en biogenese (synthese, glycosylering, mensenhandel en afbraak) eigenschappen 1. Elk van deze processen is uniek voor elk type ionenkanalen en is geoptimaliseerd om de fysiologische rol van het ionkanaal voldoen. Derhalve veranderingen in elk van deze verfijnde 'via deerfelijke of door genetische modificatie, vaak aangeduid als "channelopathy", kan schadelijk celhomeostase zijn. Het is belangrijk te benadrukken dat het leveren van de "juiste" hoeveelheid ionenkanalen aan het celoppervlak is essentieel voor celhomeostase. Zelfs kleine stijgingen (gain-of-functie) en kleine dalingen (verlies-van-functie) in ionkanaal activiteit hebben de potentie om een ernstige pathologie over een leven leiden. Defecten in het celoppervlak levering van rijpe ionkanalen is een belangrijke determinant in talrijke kanalopathieën, zoals cystische fibrose (CFTR ionkanaal) 2 en hartritmestoornissen van het lange QT syndroom vorm (cardiale kaliumkanalen) 3.
Kanalopathieën worden geassocieerd met cardiale plotselinge dood 4. De huidige wereldwijde prevalentie van alle cardiale kanalopathieën wordt gedacht dat minstens 1 te zijn: 2,000-1: 3000 per persoon 5 en zijn verantwoordelijk voor ongeveer de helft van een plotselinge aritmische hartdood cases 6. Disfunctie bij cardiale spanningsafhankelijke natrium-, kalium-, calcium- en selectieve ionenkanalen zijn bekend een belangrijke rol spelen in dit proces. De L-type Ca V 1.2-voltage-gated calcium kanaal is vereist om gesynchroniseerd hartspier contractie te starten. De cardiale L-type Ca 1,2 V kanaal is een multi-subeenheid proteïne complex samengesteld uit het porievormende Ca V α1 subeenheid en Ca Vss en Ca V α2δ1 aanvullende subeenheden 7-12. Merk op dat de volledige set van hulp- subeenheden vereist om functionele Ca V 1,2 kanalen in het plasmamembraan en dynamische interacties veroorzaken tussen deze subeenheden zijn essentieel voor de normale elektrische functie van het hart 13 te ondersteunen. Ca V ß bevordert het celoppervlak expressie van ca 1,2 V kanalen via een niet-covalente hydrofobe interactie nanomolair 14. Co-expressie van de Ca V α2δ1 subunit with Ca V ß-gebonden Ca V α1 stimuleert piekstroom uitdrukking (5 tot 10-voudig) en bevordert kanaal activering op meer negatieve spanningen. Gain-of-function mutaties van het porievormende subeenheid Ca V 1,2 zijn geassocieerd met een vorm van ventriculaire aritmieën genoemd het lange QT syndroom 15 dat samen met puntmutaties in de drie subeenheden die het L-type Ca V 1,2 kanaal geïdentificeerd in individuen die lijden aan aritmieën van de korte QT syndroom vorm 16,17. Ion kanalen zijn membraaneiwitten die kunnen worden onderzocht vanuit een biochemisch perspectief (eiwit chemie) of met behulp van elektrofysiologische instrumenten (huidige genererende machines) en vaak met behulp van deze complementaire aanpak. Elektrofysiologie, met name whole-cell patch-klemming, is een geschikte benadering voor de functie van ionkanalen 15 verhelderen maar kan niet wijzigingen in proteïne transport lossen van veranderingen in hun biofysischeeigenschappen. Eiwitchemie is echter vaak beperkt gebruik door de relatief lage expressie van grote membraaneiwitten opzichte van kleinere oplosbare eiwitten. Robuuste high-throughput methoden met behulp van fluorescentie uitlezing moeten worden ontwikkeld om specifiek ingegaan op gebreken in eiwitten biogenese veroorzaken van veranderingen in het celoppervlak expressie van ionkanalen.
Flowcytometrie is een biofysische technologie gebruikt in cel tellen, sorteren, biomarker opsporing en protein engineering 18. Wanneer een monsteroplossing van levende cellen of deeltjes cytometer in een stroom wordt geïnjecteerd, worden de cellen gerangschikt in een enkele stroom die kan worden gesondeerd door de machine detectiesysteem (figuur 1). De eerste stromingscytometer akte in 1956 19 gedetecteerd slechts één parameter maar modern flowcytometers meerdere lasers en fluorescentiedetectoren dat de detectie van meer dan 30 fluorescerende 20,21 parameters mogelijk.Filters en spiegels (emissie optiek) richt de lichtverstrooiing of TL-licht van cellen aan een elektronisch netwerk (fotodiode en detectoren), die het licht proportioneel om te zetten in de intensiteit ervan. Digitale gegevens worden geanalyseerd met gespecialiseerde software en de eerste uitgang wordt weergegeven als puntenplot 21.
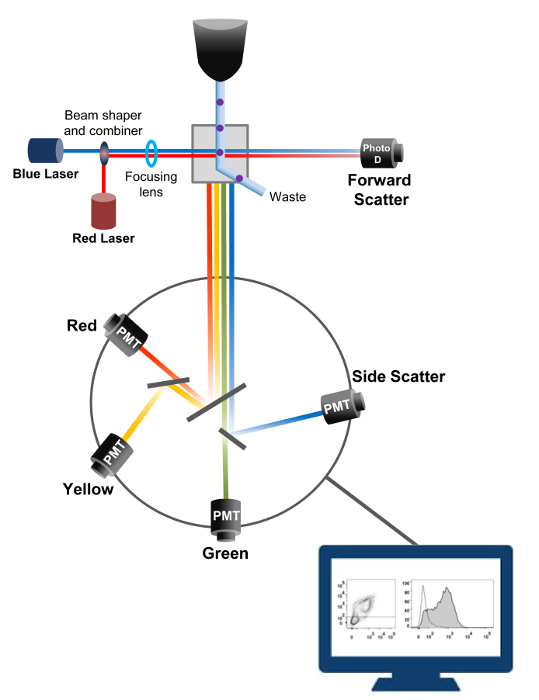
Figuur 1:. Biofysische principes flowcytometrie sorteren Single cellen worden geduwd door een sproeier onder hoge druk in een stroom omhullingfluïdum die hen beweegt op een of meerdere laser afvraagpunten. De lichtbundel wordt afgebogen door de passerende cellen en verzameld in de voorwaartse richting (Forward Scatter, FCS) licht naar een fotodiode die het licht omzet in een signaal evenredig aan de grootte van de cel. Het licht wordt ook verzameld op een 90 ° hoek met de laser pad en naar detectors (ook wel fotomultipliers (PMT)).Dit licht wordt via dichroïsche spiegels die de detectie van het signaal zijwaartse verstrooiing (SSC), die de korreligheid in de cellen weerspiegelt mogelijk, en de fluorescente emissie als opgewonden fluorochromen in de cel aanwezig zijn. Drie detectoren (groen, geel en rood) worden weergegeven met verschillende golflengte banddoorlaatfilters, waardoor de gelijktijdige detectie van verschillende fluorochromen. De verschillende signalen worden gedigitaliseerd door een externe computer en omgezet in gegevens die worden geanalyseerd om de eigenschappen van de cellen te kwantificeren. Klik hier om een grotere versie van deze figuur te zien.
De high-throughput capaciteit van flowcytometers werd benut om de relatieve membraan expressie van recombinant wild-type en de handel-deficiënte voltage-gated L-type Ca V 1.2 kanalen en bijbehorende subeenheden in levende cellen te kwantificeren. cDNA constructen coding voor de eiwitten werden dubbel gelabeld om gelijktijdig een extracellulair niet fluorescerende epitoop die door een ondoordringbare fluorescerend geconjugeerd antilichaam en een intracellulair fluorofoor die constitutief fluorescerende kan worden gedetecteerd. Zowel de extracellulaire epitoop, een extracellulaire lus van het eiwit geplaatst, en de intracellulaire fluorofoor, ingevoegd na de C-terminus, worden omgezet met het eiwit. In deze reeks experimenten werd het Ca V α2δ1 eiwit ontworpen om een extracellulaire hemagglutinine (HA) epitoop (YPYDVPDYA) gedetecteerd door een ondoordringbare FITC (fluoresceïne isothiocyanaat) expressie geconjugeerd anti-HA en mCherry de intrinsieke intracellulaire fluorofoor. De relatieve celoppervlak expressie van het mCherry-Ca V α2δ1 HA-gemerkt eiwit te bepalen, werden recombinante cellen die het fusie-eiwit na transfectie geoogst en gekleurd met FITC-geconjugeerd monoklonaal anti-HA-epitooplabel Antibody (figuur 2). FITC is een organische fluorescente verbinding die aanzienlijk kleiner is dan enzym verslaggevers en dus niet zoveel kans om te interfereren met de biologische functie. mCherry- Ca V α2δ1 HA-overexpressie in TSA-201cells, levert aanzienlijk 3-log toename van de FITC fluorescentie en fluorescentie mCherry twee-dimensionale plots 22. Aangezien de HA epitoop in het extracellulaire deel van het eiwit, de fluorescentie-intensiteit van FITC verkregen in aanwezigheid van intacte cellen weerspiegelen de relatieve index van het celoppervlak expressie van HA-gemerkt eiwit. De toegankelijkheid van de HA epitoop in de constructen systematisch gevalideerd door het FITC signaal nadat cel permeabilisatie. Deze maatregel dient ook om de genormaliseerde totale eiwitexpressie bevestigen aangezien de relatieve fluorescentie intensiteiten voor FITC in gepermeabiliseerde cellen geschat zijn kwalitatief vergelijkbaar met de relatieve fluorescentiewaarden for mCherry gemeten onder permeabel en non-gepermeabiliseerde voorwaarden 22,23. Het is belangrijk op te merken dat de intrinsieke fluorescentie spectrum wordt verschoven naar hogere waarden na permeabilisatie maar dat de enige waarde wordt gerapporteerd is de verandering in fluorescentie-intensiteit in vergelijking met de controle construct. Relatieve veranderingen in de fluorescentie-intensiteit voor de test constructen worden geschat aan de hand van de ΔMean Fluorescence Intensity (ΔMFI) waarden voor elke fluorofoor (mCherry of FITC). Experimenten zijn ontworpen om de fluorescentie-intensiteit van de testconstruct opzichte van de fluorescentie-intensiteit van de controle construct tot expressie gebracht onder dezelfde omstandigheden gemeten experimentele variaties beperken de intrinsieke fluorescentie van het fluorofoor-geconjugeerde antilichaam. Twee membraaneiwitten met succes onderzocht met behulp van deze test: de porie vormende subeenheid van het L-type spanningsafhankelijke Ca calciumkanaal V 1,2 14,22 en een andere reeksexperimenten, de extracellulaire extra Ca V α2δ1 subunit 22,23. Het volgende protocol werd toegepast om de celoppervlak expressie van het Ca V α2δ1 subunit van het cardiale L-type Ca 1,2 V kanaal onder controle omstandigheden en na mutaties die de posttranslationele modificatie van het ionkanaal te bepalen. Onder gestandaardiseerde experimentele omstandigheden het celoppervlak fluorescentie van FITC verhoogt quasi-lineair met de expressie van cDNA dat codeert voor mCherry-Ca V α2δ1 HA-eiwitten (figuur 5 van referentie 22).
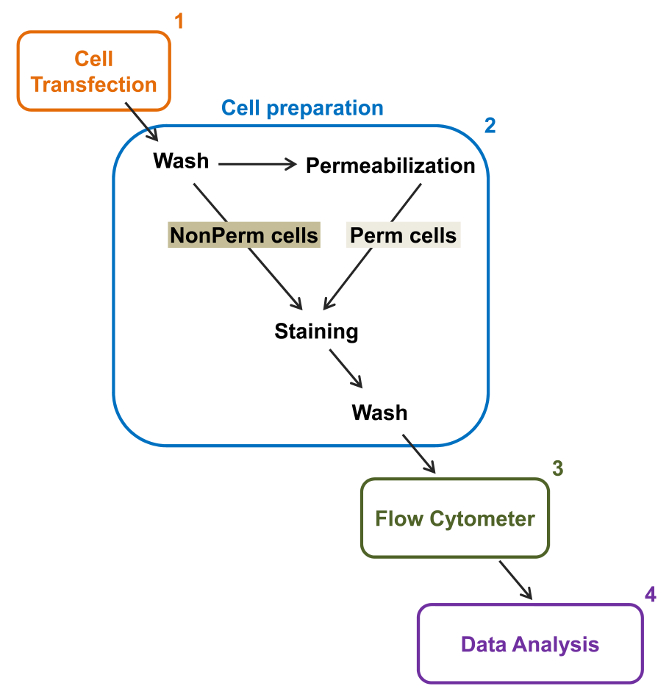
Figuur 2:. Schematische weergave van de totale en het membraan etikettering in de flowcytometrie experimentele protocol De regeling schetst enkele van de belangrijkste stappen die nodig zijn om de relatieve totale en celoppervlak expressie van recombinante ionkanalen door fl kwantificerenow cytometrie. Cellen worden getransfecteerd met de twee merktekens constructie mCherry-Ca V α2δ1-HA in TSA-201 cellen (1) en gekleurd voor of na permeabilisatie (2). Multiparameter gegevens worden verkregen in een flowcytometer (3) voor multivariate analyse (4). Klik hier om een grotere versie van deze figuur te bekijken.
