Dette papir gir en pålitelig analyse for å rapportere relativ celleoverflate-ekspresjon av membranproteiner slik som ionekanaler uttrykt i rekombinante celler ved bruk av eksisterende teknologi flowcytometri. Ionekanaler er poredannende membranproteiner som er ansvarlig for styring av elektriske signaler ved å portstyre strømmen av ioner gjennom cellemembranen. De blir klassifisert av aktiveringsmekanismen, natur, og selektiviteten til ion-arter i transitt gjennom porene hvor de er lokalisert. På de cellulære og vev nivåer, de makroskopiske ion fluks gjennom ionekanaler er et produkt av biofysiske (gating og gjennomtrengning), biokjemisk (fosforylering), og biogenesis (syntese, glykosylering, trafficking, og nedbrytning) eiendommer 1. Hver av disse prosessene er unik for hver type av ionekanaler og er optimalisert for å oppfylle den fysiologiske rollen til ionekanal. Følgelig, endringer i noen av disse finjusterte prosesser gjennom enarvet eller en genetisk modifikasjon, ofte referert til som "ionekanalsvikt", kan være skadelig for celle homeostase. Det er viktig å understreke at levere den "riktige" mengde av ionekanaler på celleoverflaten er kritisk for celle homeostase. Selv små økninger (gain-of-funksjon) og små reduksjoner (tap-av-funksjon) i ionekanal aktivitet har potensial til å forårsake en alvorlig patologi over livsløpet. Defekter i celleoverflaten levering av modne ionekanaler er en viktig determinant i en rekke kanalopatier, som cystisk fibrose (CFTR ion kanal) 2 og hjertearytmier av lang QT-syndrom form (kardiale kaliumkanaler) 3.
Kanalopatier er assosiert med hjerte plutselig død 4. Den nåværende globale utbredelsen av alle hjerte kanalopatier antas å være minst 1: 2,000-1: 3000 per individ 5 og er ansvarlig for omtrent halvparten av plutselige arytmisk hjertedød cases 6. Dysfunksjon i hjerte de spenningsstyrte natrium, kalium- og kalsium-selektive ionekanaler er kjent for å spille en nøkkelrolle i denne prosessen. Den L-type Ca V 1.2 de spenningsstyrte kalsiumkanal er nødvendig for å initiere synkronisert hjertemuskelsammentrekning. Kardiale L-type Ca V 1.2 kanal er en multi-underenhet-protein-komplekset som består av de viktigste poredannende Ca V α1-subenheten og Ca V ß og Ca V α2δ1 tilleggs subenheter 7-12. Vær oppmerksom på at full bemanning av hjelpe subenheter er nødvendig for å produsere funksjonelle Ca V 1.2 kanaler på plasmamembranen og dynamiske samspillet mellom disse underenhetene er viktig å støtte den normale elektriske funksjon av hjertet 13. Ca V ß fremmer celleoverflate-ekspresjon av Ca V 1.2 kanaler gjennom et ikke-kovalent nanomolar hydrofob interaksjon 14. Co-uttrykk for Ca V α2δ1 subenhet with Ca V ß-bundet Ca V α1 stimulerer peak strøm uttrykk (5-10 ganger) og fremmer kanal aktivering ved flere negative spenninger. Gain-av-funksjon mutasjoner av den poredannende subenhet Ca V 1.2 har vært forbundet med en form av ventrikulære arytmier som kalles forlenget QT-syndrom 15, mens en rekke av punktmutasjoner i de tre viktigste subenheter som danner L-type Ca V 1.2 kanal det er identifisert hos personer som lider av arytmier av kort QT-syndrom skjema 16,17. Ionekanaler er membranproteiner som kan undersøkes fra et biokjemisk perspektiv (protein kjemi) eller ved bruk av elektroverktøy (strømgenererende maskiner) og ofte ved hjelp av disse komplementære tilnærminger. Elektro, spesielt hel-celle patch-fastspenning, er en passende metode for å klargjøre funksjonen av ionekanaler 15, men kan ikke løse endringer i protein handel fra endringer i deres biofysiskeegenskaper. Protein kjemi har imidlertid ofte begrenset bruk på grunn av den relativt lave ekspresjon av store membranproteiner i forhold til mindre oppløselige proteiner. Robuste high-throughput metoder ved hjelp av fluorescens avlesning må utvikles for å spesielt ta opp mangler i protein biogenesis forårsaker endringer i celleoverflaten uttrykk for ionekanaler.
Flowcytometri er en biofysisk teknologi ansatt i celle telling, sortering, biomarkør oppdagelse, og protein engineering 18. Når en prøveoppløsning av levende celler eller partikler injiseres i et flowcytometer, blir cellene organisert til en enkelt strøm som kan bli probet ved maskinens deteksjonssystem (figur 1). Den første strømningscytometer instrument fremstilt i 1956 19 detektert bare en parameter, men moderne væskestrømsfotometere har flere lasere og fluorescens-detektorer som tillater påvisning av mer enn 30 fluoriserende parametere 20,21.Filtre og speil (utslipps optikk) dirigere lysspredning eller fluorescerende lys av celler til et elektronisk nettverk (fotodiode og detektorer) som konverterer lys proporsjonalt med intensiteten. Digitale data er analysert ved hjelp av spesialisert programvare og den primære effekt vises som et prikkplott 21.
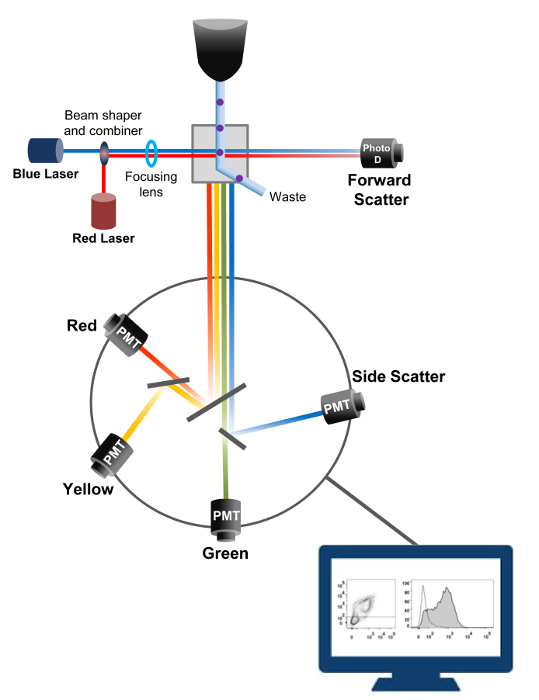
Fig. 1: Biofysiske prinsippene ved flowcytometri sortering Enkeltceller blir presset gjennom en dyse under høyt trykk i en strøm av skjermfluid som beveger dem på tvers av ett eller flere laserutspørrings poeng. Lysstrålen avbøyes av de passerende celler, og lyset samles i fremoverretningen (fremover diffusjon, FCS) blir sendt til en fotodiode som omdanner lys til et signal proporsjonalt med størrelsen av cellen. Lyset er også samlet i en 90 ° vinkel til laser banen og sendt til detektorene (også kalt fotomultiplikatorer (PMT)).Dette lyset ledes gjennom dikroiske speil som tillater påvisning av sidespredningssignalet (SSC), som gjenspeiler den detaljnivået i cellene, og de fluorescerende utslipp hvis eksiterte fluorokromer er til stede i cellen. Tre detektorer (grønn, gul, og rød) er representert med ulike bølgelengde båndpassfiltre, slik at den samtidig påvisning av ulike fluorokromer. De forskjellige signalene digitalisert av en ekstern datamaskin og konvertert til data som vil bli analysert for å kvantifisere egenskapene til cellene. Klikk her for å se en større versjon av dette tallet.
Den high-throughput kapasitet på flowcytometere ble utnyttet til å kvantifisere den relative membran uttrykk av rekombinant villtype og trafficking-mangelfull spennings gated L-type Ca V 1.2 kanaler og tilhørende underenhetene i levende celler. cDNA konstruerer coding for proteinene var dobbelt merket for samtidig å bære en ekstracellulær ikke-fluoriserende epitop som kan oppdages ved hjelp av en ugjennomtrengelig fluorescerende antistoff og et intracellulært fluorofor som er konstitutivt fluorescerende. Både den ekstracellulære epitop, innføres i et ekstracellulært løkke av proteinet, og den intracellulære fluorofor, innføres etter den C-terminale enden, blir omregnet med proteinet. I denne serien av forsøk ble Ca V α2δ1 proteinet konstruert for å uttrykke et ekstracellulært hemagglutinin (HA) epitop (YPYDVPDYA) som detekteres ved hjelp av en ugjennomtrengelig FITC (fluoresceinisotiocyanat) -konjugert anti-HA og mCherry som den indre intracellulære fluoroforen. For å bestemme den relative celleoverflate-ekspresjon nivået av mCherry-Ca V α2δ1 HA-kodede protein, ble rekombinante celler som uttrykker fusjonsproteinet høstet etter transfeksjon, og farget med FITC-konjugert mus monoklonalt anti-HA-epitop tag antibody (figur 2). FITC er en organisk fluorescerende stoff som er betydelig mindre enn enzym journalister og derfor ikke så sannsynlig å påvirke biologisk funksjon. mCherry- Ca V α2δ1-HA overexpressed i TSA-201cells, produserer en betydelig tre-log økning i FITC fluorescens og mCherry fluorescens på todimensjonale plott 22. Gitt at HA-epitop er lokalisert i den ekstracellulære del av proteinet, fluorescensintensiteten for FITC oppnådd i nærvær av intakte celler gjenspeiler den relative indeksen til celleoverflate-ekspresjon av HA-kodede protein. Tilgjengeligheten av HA-epitop i konstruksjonene er systematisk validert ved å måle FITC signal etter celle permeabilization. Dette tiltaket tjener også til å underbygge den normaliserte totale protein ekspresjon, siden de relative intensiteter for FITC fluorescens estimert i permeabiliserte celler er kvalitativt sammenlignbare med de relative fluorescens-verdier for mCherry målt under permeabilized og ikke-permeabilized forhold 22,23. Det er viktig å merke seg at den indre fluorescens-spekteret er forskjøvet mot høyere verdier etter permeabiliseringen men at den eneste verdi som rapporteres er endringen i fluorescens-intensitet sammenlignet med kontroll-konstruksjonen. Relative endringer i fluorescens intensitet for test konstruksjoner blir estimert ved hjelp av ΔMean fluorescensintensitet (ΔMFI) verdier for hver fluorophore (mCherry eller FITC). Eksperimenter er utformet for å måle fluorescensintensiteten av test konstruksjonen i forhold til fluorescensintensiteten av styre konstruksjonen uttrykt under de samme betingelser for å begrense eksperimentelle variasjoner i den indre fluorescens av det fluorofor-konjugert antistoff. To membranproteiner ble med hell undersøkt ved hjelp av denne analysen: den poredannende subenhet av L-typen de spenningsstyrte kalsiumkanal Ca V 1,2 14,22 og i en annen serieeksperimenter, den ekstracellulære hjelpe Ca V α2δ1 subenhet 22,23. Den følgende protokoll ble anvendt for å bestemme celleoverflate-ekspresjon av Ca V α2δ1 subenheten til hjerte L-type Ca V 1,2 kanalen under kontrollbetingelser, og etter mutasjoner som påvirker posttranslasjonell modifikasjon av ionekanal. Under standardiserte eksperimentelle betingelser, celleoverflaten fluorescensen av FITC øker kvasi-lineært med ekspresjon av cDNA som koder for de mCherry-Ca V α2δ1-HA-proteiner (figur 5 fra referanse 22).
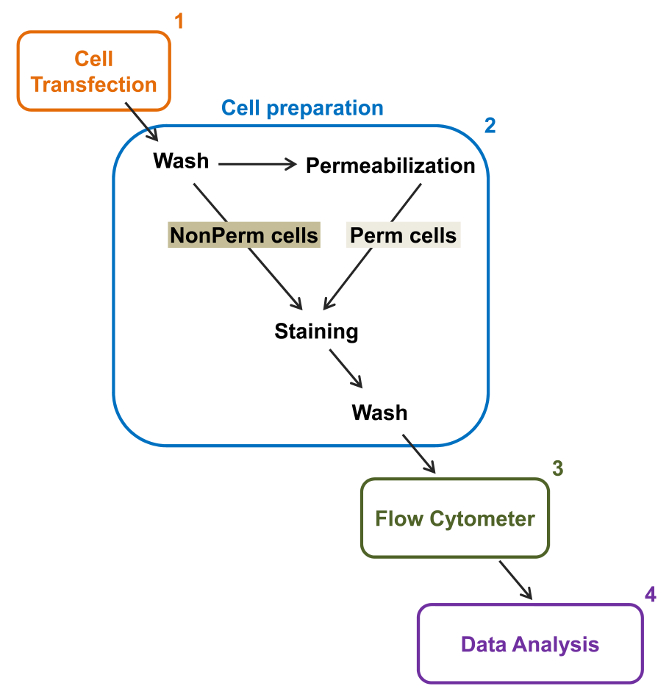
Fig. 2: Skjematisk representasjon av total og membranmerking i flowcytometri forsøksprotokoll Ordningen skisserer noen av de hovedtrinn som er nødvendige for å kvantifisere den relative totale og celleoverflate-ekspresjon av rekombinante ionekanaler etter flow cytometri. Cellene transfektert med dobbel-taggede konstruksjon mCherry-Ca V α2δ1-HA i TSA-201 celler (1) og farget før eller etter permeabilization (2). Funksjoner med flere data er kjøpt i et flowcytometer (3) for multivariat analyse (4). Klikk her for å se en større versjon av dette tallet.
