Este documento proporciona un ensayo fiable para informar de la superficie celular de expresión relativa de las proteínas de membrana como canales iónicos expresados en las células recombinantes utilizando la tecnología de citometría de flujo existente. Los canales iónicos son proteínas de membrana formadores de poros que son responsables del control de señales eléctricas por gating el flujo de iones a través de la membrana celular. Se clasifican por el mecanismo de activación, la naturaleza, y la selectividad de las especies de iones que transitan a través del poro donde se localizan. En los niveles celulares y tisulares, los flujos de iones a través de los canales iónicos macroscópicas son el producto de las propiedades biofísicas 1 (gating y la permeabilidad), bioquímica (fosforilación), y la biogénesis (síntesis, glicosilación, el tráfico y la degradación). Cada uno de estos procesos es única para cada tipo de canales de iones y está optimizado para cumplir el papel fisiológico del canal de iones. En consecuencia, las alteraciones en cualquiera de estos procesos-finas sintonizado a través de unaheredada o una modificación genética, a menudo referido como "canalopatía", puede ser perjudicial para la homeostasis celular. Es importante hacer hincapié en que la entrega de la cantidad "correcta" de los canales iónicos en la superficie celular es crítica para la homeostasis celular. Incluso pequeños incrementos (ganancia de función) y disminuciones pequeñas (pérdida de la función) en la actividad de los canales iónicos tienen el potencial de causar una patología grave durante toda la vida. Los defectos en la entrega de la superficie celular de los canales iónicos maduro es un determinante importante en numerosos canalopatías, como la fibrosis quística (CFTR canal iónico) 2 y arritmias cardiacas de forma síndrome de QT largo (canales de potasio cardíacos) 3.
Canalopatías se asocian con la muerte súbita cardiaca 4. La prevalencia actual en todo el mundo de todas las canalopatías cardiacas se piensa que es al menos 1: 2,000-1: 3,000 por persona 5 y son responsables de cerca de la mitad de la súbita arrítmica ca muerte cardíacases 6. Disfunción cardíaca en sodio dependientes de voltaje, potasio, calcio y los canales iónicos selectivos se sabe que juegan un papel clave en este proceso. Se requiere que el 1.2 canales de calcio dependientes de voltaje tipo L Ca V para iniciar la contracción del músculo cardíaco sincronizado. La cardiaca de tipo L Ca V 1.2 canales es un complejo de proteínas de múltiples subunidades compuesto por el principal Ca subunidad α1 V de formación de poros y Ca V SS y Ca V α2δ1 subunidades auxiliares 7-12. Tenga en cuenta que se requiere el complemento completo de subunidades auxiliares para producir funcionales Ca V 1.2 canales en la membrana plasmática y las interacciones dinámicas entre estas subunidades son esenciales para apoyar la función eléctrica normal del corazón 13. Ca V ß promueve la expresión de la superficie celular de Ca V 1.2 canales a través de un nanomolar de interacción hidrofóbica no covalente 14. La co-expresión de la subunidad V wi α2δ1 Caunido a la ß TH Ca V V α1 Ca estimula la expresión corriente de pico (de 5 a 10 veces) y promueve la activación del canal a voltajes más negativos. Ganancia de función de las mutaciones de la subunidad formadora de poros Ca V 1.2 se han asociado con una forma de arritmias ventriculares llamado el síndrome de QT largo 15 mientras que una serie de mutaciones puntuales en las tres subunidades principales que forman el tipo L Ca V 1.2 canales han sido identificados en sujetos que sufren de arritmias de forma síndrome de QT corto 16,17. Los canales iónicos son proteínas de membrana que pueden ser investigados desde una perspectiva bioquímica (la química de proteínas) o utilizando herramientas electrofisiológicos (máquinas generadoras de corriente) y, a menudo utilizando estos enfoques complementarios. Electrofisiología, en particular de células enteras patch-sujeción, es un enfoque adecuado para dilucidar la función de los canales iónicos 15 pero no puede resolver modificaciones en el tráfico de proteínas a partir de los cambios en su biofísicopropiedades. química de proteínas ha, sin embargo, a menudo limitado el uso debido a la relativamente baja expresión de grandes proteínas de la membrana en relación con las proteínas solubles más pequeños. Robustos métodos de alto rendimiento utilizando la lectura de fluorescencia se deben desarrollar con el fin de abordar específicamente los defectos de la biogénesis de proteínas que causan cambios en la expresión de la superficie celular de los canales iónicos.
La citometría de flujo es una tecnología biofísico empleada en el recuento de células, la clasificación, la detección de biomarcadores y la ingeniería de proteínas 18. Cuando se inyecta una solución de muestra de células vivas o partículas en un citómetro de flujo, las células se ordenan en una única corriente que se puede probar por el sistema de detección de la máquina (Figura 1). La primera citómetro de flujo instrumento producido en 1956 19 detectado sólo un parámetro pero citómetros de flujo modernos tienen múltiples láseres y detectores de fluorescencia que permiten la detección de más de 30 parámetros fluorescentes 20,21.Filtros y espejos (óptica de emisión) dirigen la dispersión de la luz o luz fluorescente de las células a una red electrónica (fotodiodo y detectores) que convierten la luz en proporción a su intensidad. Los datos digitales se analizan usando un software especializado y la salida principal se muestra como un gráfico de puntos 21.
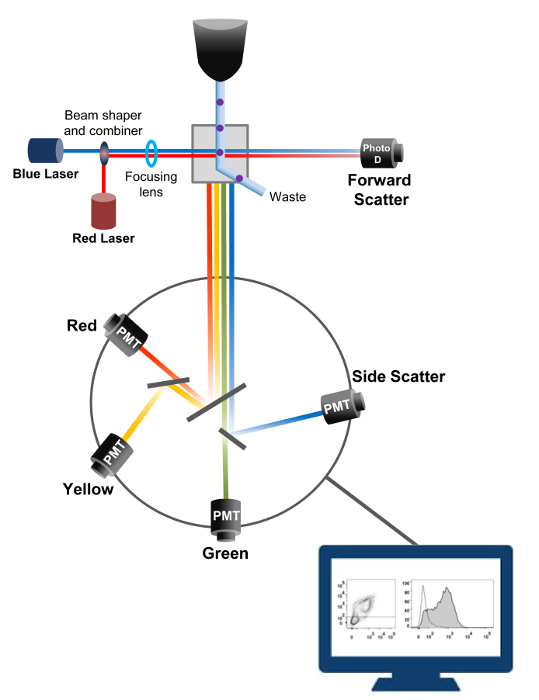
Figura 1:. Principios biofísicos de citometría de flujo clasificar las células individuales son empujados a través de una boquilla a alta presión dentro de una corriente de fluido de funda que les mueve a través de uno o más puntos de interrogación láser. El haz de luz es desviado por las células que pasan y la luz recogida en la dirección hacia adelante (Forward Scatter, FCS) se envía a un fotodiodo que convierte la luz en una señal proporcional al tamaño de la célula. La luz también se recoge en un ángulo de 90 ° a la trayectoria de láser y se envía a detectores (también llamados fotomultiplicadores (PMT)).Esta luz se dirige a través de espejos dicroicos que permiten la detección de la señal de dispersión lateral (SSC), que refleja la granularidad dentro de las células, y las emisiones fluorescentes si fluorocromos excitados están presentes en la célula. Tres detectores (verde, amarillo y rojo) se representan con diferentes filtros de paso de banda de longitud de onda, lo que permite la detección simultánea de diferentes fluorocromos. Las diferentes señales son digitalizadas por un ordenador externo y se convierten en datos que serán analizados para cuantificar las características de las células. Por favor, haga clic aquí para ver una versión más grande de esta figura.
La capacidad de alto rendimiento de citómetros de flujo fue explotada para cuantificar la expresión en la membrana relativa de tipo salvaje recombinante y la trata con deficiencia de voltaje de tipo L Ca V 1.2 canales y subunidades asociadas en las células vivas. construcciones de ADNc coding para las proteínas se marcaron doblemente para llevar simultáneamente un epítopo no fluorescente extracelular que puede ser detectado por un anticuerpo conjugado fluorescente impermeable y un fluoróforo intracelular que es constitutivamente fluorescente. Tanto el epítopo extracelular, insertado en un bucle extracelular de la proteína, y el fluoróforo intracelular, insertado después de la C-terminal, se traducen con la proteína. En esta serie de experimentos, la proteína V α2δ1 Ca fue diseñado para expresar una hemaglutinina extracelular (HA) epítopo (YPYDVPDYA) detectada por un impermeable FITC (isotiocianato de fluoresceína) conjugada anti-HA y mCherry como el fluoróforo intracelular intrínseco. Para determinar el nivel de expresión de la superficie celular relativa de la V α2δ1 mCherry-Ca proteína etiquetada con HA, se recogieron las células recombinantes que expresan la proteína de fusión después de la transfección, y se tiñeron con el anticuerpo monoclonal de ratón anti-HA antibod etiqueta de epítopo conjugado con FITCy (Figura 2). FITC es un compuesto fluorescente orgánico que es considerablemente más pequeño que los reporteros de la enzima y por lo tanto no es tan probable que interfiera con la función biológica. mCherry- Ca V α2δ1-HA sobreexpresa en TSA-201cells, produce un aumento de 3 registro significativo de la fluorescencia FITC y la fluorescencia mCherry en gráficos bidimensionales 22. Dado que el epítopo HA se encuentra en la porción extracelular de la proteína, la intensidad de fluorescencia de FITC obtiene en presencia de células intactas reflejan el índice relativo de la expresión de la superficie celular de la proteína etiquetada con HA. La accesibilidad del epítopo HA en las construcciones se valida sistemáticamente mediante la medición de la señal de FITC después de la permeabilización celular. Esta medida también sirve para corroborar la expresión de la proteína total normalizada ya que las intensidades relativas de fluorescencia de FITC estimaron en células permeabilizadas son cualitativamente comparables con los valores de fluorescencia relativa FOr mCherry mide bajo permeabilizadas y no permeabilizadas condiciones 22,23. Es importante tener en cuenta que el espectro de fluorescencia intrínseca se desplaza hacia valores más altos después de la permeabilización, pero que el único valor que se informa es el cambio en la intensidad de fluorescencia en comparación con la construcción de control. Los cambios relativos en la intensidad de fluorescencia de las construcciones de ensayo se calculan utilizando la intensidad de fluorescencia ΔMean (ΔMFI) los valores para cada fluoróforo (mCherry o FITC). Los experimentos están diseñados para medir la intensidad de fluorescencia de la construcción de ensayo con respecto a la intensidad de fluorescencia de la construcción de control expresado en las mismas condiciones para limitar las variaciones experimentales en la fluorescencia intrínseca del anticuerpo fluoróforo conjugado. Dos proteínas de la membrana fueron estudiados con éxito utilizando este ensayo: la subunidad formadora de poros del canal de calcio dependiente de voltaje de tipo L Ca V 1.2 14,22 y en una serie diferente deexperimentos, el auxiliar extracelular Ca V α2δ1 subunidad 22,23. Se usó el siguiente protocolo para determinar la expresión de la superficie celular de la Ca V α2δ1 subunidad de la cardíaco L-tipo Ca V 1.2 canal en condiciones de control y después de las mutaciones que afectan a la modificación postraduccional del canal iónico. En condiciones experimentales estandarizadas, la fluorescencia de la superficie celular de FITC aumenta casi linealmente con la expresión de ADNc que codifica para las proteínas α2δ1-HA mCherry-Ca V (Figura 5 de la referencia 22).
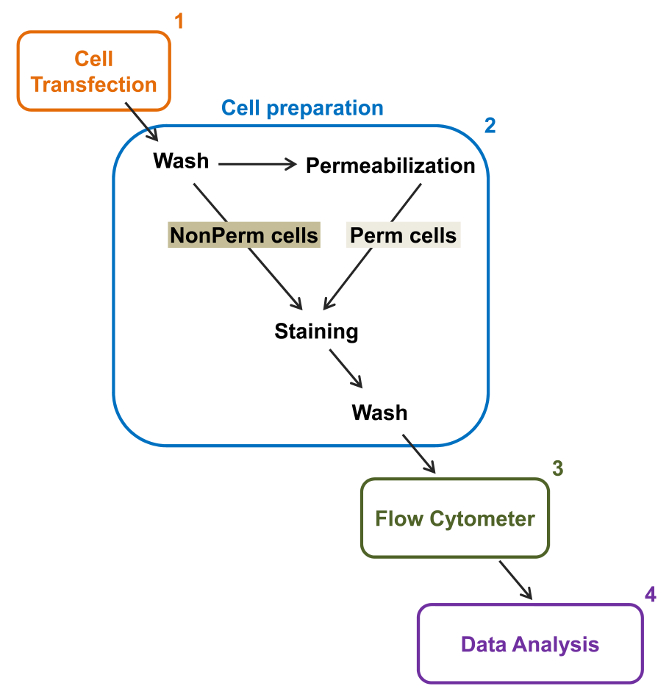
Figura 2:. Representación esquemática de etiquetado total y membrana en la citometría de flujo protocolo experimental El esquema se describen algunos de los principales pasos necesarios para cuantificar la expresión total y superficie celular relativa de los canales iónicos recombinantes por flflujo citometría. Las células se transfectaron con la construcción de doble etiquetado mCherry-Ca V α2δ1-HA en TSA-201 células (1) y se tiñeron antes o después de la permeabilización (2). Multiparamétricos datos se adquieren en un citómetro de flujo (3) para el análisis multivariado (4). Haga clic aquí para ver una versión más grande de esta figura.
