تحرير الجينات الدقيقة تشكل حجر الزاوية للتطورات الطبية الحيوية الرئيسية التي تنطوي على وضع استراتيجيات جديدة للتصدي للأمراض الوراثية. في طليعة تكنولوجيا الجينات–تحرير بالطريقة التي تعتمد على استخدام جلوستيريد rاجولارلي–أنانتيرسباسيد sهورت فاليندروميك صابيتس (كريسبر)/نظام Cas9 التي تم تحديدها في البداية كمكون للبكتيريا مناعة ضد الغزو المواد الجينية الفيروسية (إعادة النظر في المراجع1،2). ميزة كبيرة للنظام كريسبر/Cas9 على غيرها من أدوات تحرير الجينات، مثل نوكلياسيس إصبع الزنك (زفنس) والنسخ المستجيب المنشط–مثل نوكلياسيس (تالينس) (واستعرضت في مرجع3)، هي البساطة النسبية لتصميم بلازميد و بناء مكونات كريسبر – ميزة التي لديه بدعم التوسع في تحرير الجينات من عدد قليل من المختبرات المتخصصة إلى مجتمع بحث أوسع كثيرا. بالإضافة إلى ذلك، غذت بساطة البرمجة كريسبر/Cas9 وقدرته على الاعتراف بهدف متعدد زيادة شعبيتها كتكنولوجيا فعالة من حيث التكلفة وسهلة الاستخدام. بين الأساليب المختلفة المتاحة للباحثين لتقديم هذه المكونات تحرير الجينات للخلايا، يبقى النواقل الفيروسية بكثير النظام الأكثر شعبية وأكثر كفاءة.
ناقلات لينتيفيرال (LVs) ظهرت كوسيلة الاختيار لتقديم عناصر كريسبر/Cas9 النظام في فيفو لتطبيقات متنوعة4،5،،من67. جعل العديد من الميزات الرئيسية LVs خياراً شعبيا لهذه العملية بما في ذلك قدرتها على إصابة الفاصل وعدم تقسيم الخلايا، الاستمناع منخفضة، والسمية الخلوية الحد الأدنى (استعرض في المرجع8). نتيجة لذلك استخدمت العلاج الجيني بوساطة LV في علاجات للأمراض المعدية، مثل فيروس نقص المناعة البشرية-1 و HBV هامبورغ-1، وكذلك في تصحيح العيوب الكامنة وراء الأمراض الوراثية البشرية، مثل التليف الكيسي والأجسام القريبة من الأرض والأوعية الدموية البقعي 4 , 5 , 7 , 9 , 10 , 11-وعلاوة على ذلك، LVs قد عدلت فعلياً تنفيذ تحرير متعدد الجينات في المكاني الجينوم متميزة باستخدام نظام ناقل واحد12.
ومع ذلك، الخاصية المتأصلة من LVs الاندماج في جينوم مضيف يمكن مطفرة وغالباً ما يعوق فائدتها كوسائل إيصال التحوير، لا سيما في ظروف سريرية. وعلاوة على ذلك، منذ LVs ستابلي المتكاملة أعرب عن المتسلسلات في مستويات عالية على نحو مستدام، وهذا النظام غير ملائمة لإيصال الجينات تحرير مكونات مثل كريسبر/Cas9؛ أوفيريكسبريسيون Cas9–دليل الحمض النووي الريبي (جرنا)، والبروتينات مشابهة مثل زفنس، ترتبط بمستويات مرتفعة من الآثار قبالة المستهدفة، والتي تشمل الطفرات غير مرغوب فيها13،14،،من1516 , 17 ويمكن أن يحتمل أن تعزز سيتوتوكسيسيتي18. ولذلك، لتحقيق دقة تحرير الجينات مع الحد الأدنى من الآثار خارج الهدف، يتحتم لتصميم النظم التي تسمح لتعبير الجينات تحرير مكونات عابرة.
وفي السنوات الأخيرة، وضعت مجموعة متنوعة من منصات التسليم لعابر إكسبريس كريسبر/Cas9 في الخلايا16،19،،من2021 (استعرض في المرجع22). وتشمل هذه الأساليب التي تعتمد على مباشرة إدخال Cas9 المنقي جنبا إلى جنب مع الكشف الدليل المناسب في الخلايا، والتي تبين أن يكون أكثر فعالية في تحرير الجينات المستهدفة بالمقارنة بوساطة بلازميد تعداء16. وقد أثبتت الدراسات أن ريبونوكليوبروتين (رنب) دليل مجمعات تتكون من الحمض النووي الريبي/Cas9 الجزيئات سرعة تسليم بعد وساطة الانقسام الحمض النووي في أهدافها، مما يدل على أن التعبير القصير الأجل لهذه المكونات تكفي لتحقيق قوة الجينات تحرير16. ومن المتصور عدم إدماج أنظمة النواقل الفيروسية مثل النواقل الفيروسية المرتبطة بالغدة (أعفس) يمكن أن توفر بديلاً مجديا لتسليم الآلات تحرير الجينات إلى الخلايا. ولسوء الحظ، كابسيدس إف تمتلك قدرة التعبئة أقل بكثير مما LVs (< 5 كيلو بايت)، مما يعوق بشدة قدرتها على حزمة أدوات كريسبر متعدد العناصر داخل ناقل واحد (استعرض في المرجع8). تجدر الإشارة إلى أن إضافة المركبات التي تمنع هيستون ديسيتيلاسيس (مثلاً، بوتيراتي الصوديوم23) أو تعيق دورة الخلية (مثلاً، الكافيين24) أظهرت زيادة التتر لينتيفيرال. وعلى الرغم من التقدم الذي أحرز مؤخرا، ما زال يعرقل النظم تعبير عابر التي وضعت حتى الآن من أوجه قصور عديدة، مثل انخفاض كفاءة الإنتاج، مما يؤدي إلى انخفاض الفيروسية التتر، وكفاءة توصيل منخفض من الفيروسات التي تم إنشاؤها من خلال وهذا النهج25.
ناقلات تفتقر إلى إينتيجراسي لينتيفيرال (إيدلفس) تمثل تقدما كبيرا في تطوير مركبات إيصال الجينات، كما أنها تجمع بين القدرة على التعبئة والتغليف من LVs مع فائدة إضافية للصيانة مثل إف ابيسومال في الخلايا. تساعد هذه الميزات إيدلفس إلى حد كبير الالتفاف حول القضايا الرئيسية المرتبطة بإدماج ناقلات، overexpression المقارنة المستمرة يحتمل أن تكون عناصر سمية جينية والتكامل بوساطة المحدثة للطفرات. قد تجلى سابقا أنه يمكن تعديل إيدلفس بنجاح لتعزيز التعبير الجيني ابيسومال26،27. فيما يتعلق بالتسليم بوساطة إيدلف كريسبر/Cas9، التتر الإنتاج المنخفض والتعبير أدنى الجينومات تنقلها ابيسومي بالنسبة لأنظمة لينتيفيرال إينتيجراسي-يتقن يحد من فائدتها كأدوات حسن النية لتقديم التحرير الجينوم بنيات المحورة وراثيا. نحن مؤخرا أظهرت أن التعبير التحوير والتتر الفيروسية المرتبطة بإنتاج إيدلف تتعزز كثيرا بإدراج مواقع لعامل النسخ حزمة الخدمة Sp1 ضمن كاسيت التعبير الفيروسية28الربط. قوة دعم إيدلفس تم التعديل الجيني بوساطة كريسبر التحرير سواء في المختبر (في الخلايا HEK-293T) و في فيفو (في الدماغ بعد الانقسامية الخلايا العصبية)، في حين حمل الطفرات خارج الهدف الحد الأدنى مقارنة بالمقابلة إيكلف بوساطة نظم28. عموما، قمنا بتطوير رواية، أدوات كريسبر المدمجة، والكل في واحد على منصة إيدلف وأوجز مختلف مزايا استخدام وسيلة إيصال لتحرير الجينات المحسنة.
هنا، بروتوكول إنتاج نظام إيدلف-كريسبر/Cas9 وصف، بما في ذلك الخطوات المختلفة التي تنطوي عليها في الجمعية، وتنقية، والتركيز، والمعايرة إيدلفس، فضلا عن استراتيجيات للتحقق من فعالية الجينات تحرير هذه العوامل الناقلة للمرض. هذا البروتوكول هو قابلة بسهولة لتلبية احتياجات مختلفة من المحققين وهو مصمم بنجاح إنشاء ناقلات LV مع التتر في النطاق من 1 × 1010 وحدات (تو) ترانسدوسينج/مل. يمكن أن تستخدم ناقلات التي تم إنشاؤها من خلال هذا البروتوكول لكفاءة تصيب العديد من أنواع الخلايا المختلفة، بما في ذلك صعوبة ترانسدوسي الخلايا الجذعية الجنينية، الخلايا المكونة للدم (خلايا تي والضامة)، ومثقف و في فيفو– حقن الخلايا العصبية. وعلاوة على ذلك، أن البروتوكول أيضا مناسبة تماما لإنتاج ناقلات لينتيفيرال إينتيجراسي المختصة بكميات مماثلة.
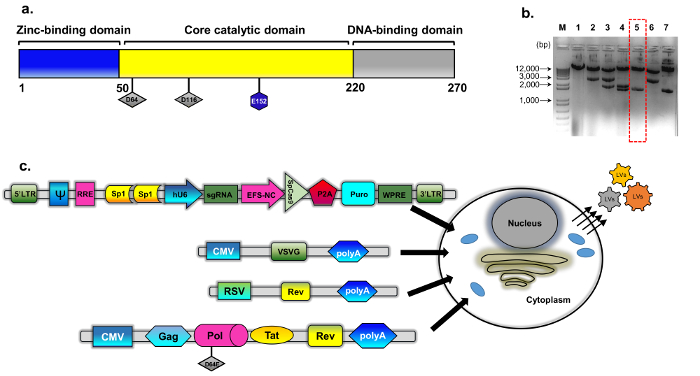
رقم 1: التعبئة والتغليف إيدلف. (أ) التخطيطي من البروتين integrase البرية نوع (ب) بلازميد معدلة مستمد من psPAX2 (راجع أساليب البناء بلازميد للحصول على التفاصيل). الممثل [اغروس] هلام صورة استنساخ فرزهم لاستنساخ integrase المتحولة. تم تحليل عينات الحمض النووي التي أعدت باستخدام مجموعة مصغرة عزل الحمض النووي بلازميد قياسية بالهضم مع اكورف وسفي. كذلك تحققت التسلسل المباشر (سانجر) لاستبدال D64E في INTاستنساخ يهضم بشكل صحيح (رقم 5، مربع أحمر متقطع). كان اسمه كاسيت التغليف تفتقر إلى إينتيجراسي pBK43. (ج) يعمل التخطيطي بروتوكول تعداء عابرة لتوليد نواقل إيدلف-كريسبر/Cas9، تظهر الخلايا 293T transfected مع منظمة-ز، والتغليف، والتحوير أشرطة الكاسيت (بلازميد الكل في واحد حزمة الخدمة Sp1-كريسبر/Cas9). الجسيمات الفيروسية أن برعم خارجاً من غشاء الخلية تحتوي على كامل طول الجيش الملكي النيبالي لمكافحة ناقلات (معبراً عنها من الكاسيت التحوير). استخدمت الجيل الثاني من نظام إيدلف والتغليف، والتي تشمل البروتينات التنظيمية تأت والقس Rev التعبير هو تكميل من كاسيت منفصل (RSV-القس-بلازميد). أبريف: لتر-منذ فترة طويلة–المحطة الطرفية فيروس التهاب الفم الحويصلي تكرار، منظمة-ز، ز-البروتين، المروج الفيروس المضخم للخلايا-بكمف؛ ساركومه راوس المروج فيروس (RSV)؛ رر-(رؤ استجابة عنصر). العناصر التنظيمية الأخرى في كاسيت التعبير تشمل مواقع Sp1-الربط، رد القس العنصر (رر)، وودتشوك التهاب الكبد الوبائي فيروس بوسترانسكريبشونال التنظيمية عنصر (وبري)، مروج 1α عامل استطالة أساسية (EFS-NC)، ناقل التعبئة والتغليف عنصر ψ (psi) والبشرية الفيروس المضخم للخلايا (hCMV) مروج ومروج U6 البشرية. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم-
