Gène précis édition constitue la pierre angulaire des grandes avancées biomédicales qui impliquent le développement de nouvelles stratégies pour lutter contre les maladies génétiques. À l’avant-garde des technologies de modification génétique est la méthode en s’appuyant sur l’utilisation de la clustered regularly –jenterspaced short palindromic repeats (CRISPR) / système de Cas9 qui a été initialement identifié dans le cadre de l’immunité contre l’invasion virale matériel génétique (révisé dans les références1,2) bactérienne. Un avantage majeur du système CRISPR/Cas9 autres gène-montage des outils, tels que les nucléases de doigt de zinc (ZFNs) et les nucléases d’effecteur comme activateur de transcription (TAPS) (révisés en référence3), est la relative simplicité de conception de plasmide et construction des composants de CRISPR — une caractéristique qui a alimenté l’expansion du gène-montage de quelques laboratoires spécialisés à une communauté de recherche beaucoup plus large. En outre, la simplicité de programmation CRISPR/Cas9 et sa capacité de reconnaissance de cible multiplexé ont alimenté plus sa popularité comme une technologie rentable et facile à utiliser. Parmi les diverses méthodes disponibles aux chercheurs de livrer ces composantes de l’édition de gène aux cellules, vecteurs viraux demeurent de loin le plus populaire et efficace système.
Vecteurs LENTIVIRAUX (LVs) sont apparus comme le véhicule de choix pour fournir les composants de CRISPR/Cas9 système in vivo pour diverses applications4,5,6,7. Les principales fonctionnalités font LVs un choix populaire pour ce processus, y compris leur capacité à infecter des cellules en division et non de division, faible immunogénicité tant minime toxicité cellulaire (revu en référence8). Ainsi, la thérapie génique induite par le LV a été employée dans le traitement de maladies infectieuses comme le VIH-1 et HSV-1, HBV, ainsi que dans la correction des défauts sous-jacents des maladies héréditaires humaines, telles que la fibrose kystique et la dégénérescence maculaire neo-vasculaire 4 , 5 , 7 , 9 , 10 , 11. par ailleurs, LVs ont été effectivement modifiées pour effectuer des retouches de gène multiplex locus génomiques distincts à l’aide d’un vecteur unique système12.
Cependant, la propriété inhérente de LVs à intégrer dans le génome hôte peut être mutagène et handicape souvent leur utilité comme vecteurs de transgène, notamment dans les milieux cliniques. En outre, depuis intégrée de façon stable LVs expriment leur transgènes durablement élevés, ce système est inadapté pour la livraison de gène-montage de composants tels que les CRISPR/Cas9 ; la surexpression de l’ARN Cas9-guide (gRNA) et des protéines semblables tels que ZFNs, sont associées à des niveaux élevés d’effets hors cible, incluent des mutations indésirables13,14,15,16 , 17 et peut potentiellement améliorer la cytotoxicité18. Par conséquent, pour atteindre précis édition de gène avec des effets minimes hors cible, il est impératif de concevoir des systèmes qui permettent l’expression transitoire du gène montage des composants.
Ces dernières années, une variété de plates-formes de diffusion ont été développés pour exprimer transitoirement CRISPR/Cas9 dans cellules16,19,20,21 (revu en référence22). Il s’agit de méthodes qui dépendent directement introduire Cas9 purifiée ainsi que le guide approprié ARN dans des cellules, qui s’est avéré plus efficace à l’édition de gène ciblé par rapport à médiation plasmidique transfection16. Des études ont démontré que la ribonucléoprotéine (RNP) complexes comprenant guident RNA/Cas9 particules sont rapidement remis après médiation de clivage de l’ADN à leurs cibles, ce qui suggère que l’expression à court terme de ces composants est suffisante pour atteindre gène robuste édition16. En théorie, sans intégration des plates-formes de vecteurs viraux tels que les vecteurs viraux adéno-associés (AAVs) peuvent fournir une alternative viable pour livrer les machines édition de gène aux cellules. Malheureusement, les capsides AAV possèdent significativement plus faible capacité de l’emballage que LVs (< 5 Ko), qui entrave gravement leur aptitude pour empaqueter le toolkit CRISPR multicomposants au sein d’un unique vecteur (revu en référence8). Il est à noter qu’ajout de composés qui inhibent les histones désacétylases (p. ex., le butyrate de sodium23) ou entraver le cycle cellulaire (p. ex., caféine24) ont été montré pour augmenter des titres des gènes. Malgré les progrès récents, les systèmes d’expression transitoire développés jusque sont encore entravés par plusieurs défauts, tels que l’efficacité de la production plus faible, conduisant à des titres viraux réduits et efficacité faible transduction des vicitimes du virus ces approches25.
Vecteurs LENTIVIRAUX intégrase déficient (IDLVs) représentent une avancée majeure dans le développement de véhicules de livraison-gène, car ils combinent la capacité de l’emballage de LVs avec l’avantage supplémentaire d’entretien épisomiques AAV-comme dans les cellules. Ces fonctionnalités permettent IDLVs de contourner en grande partie les principaux enjeux rattachés à l’intégration des vecteurs, surexpression continue vis-àvis des éléments potentiellement génotoxiques et mutagénicité induite par l’intégration. Il a été précédemment démontré que IDLVs peuvent être modifiées avec succès pour améliorer l’expression de gène épisomiques26,27. En ce qui concerne la livraison de médiation jusque CRISPR/Cas9, titres de production faible et plus faible expression des génomes épisomes-la charge par rapport aux systèmes des gènes intégrase-proficient limite leur utilité comme outils de bona fide pour la livraison du génome-montage constructions transgéniques. Nous avons récemment démontré que transgene expression tant des titres viraux associés à production jusque sont considérablement renforcés par l’inclusion de sites pour le facteur de transcription Sp1 dans l’expression virale cassette28de liaison. Les IDLVs mis à jour le fermement appuyé gènes CRISPR édition aussi bien in vitro (dans les cellules HEK-293 t) et in vivo (dans les neurones du cerveau post-mitotiques), tout en induisant des mutations hors cible minimales par rapport à la médiation ICLV correspondant 28de systèmes. Dans l’ensemble, nous avons développé un roman, compact, tout-en-un toolkit CRISPR exploite une plateforme jusqu’et décrit les divers avantages d’utiliser un tel véhicule de livraison pour l’édition de gène amélioré.
Ici, le protocole de production du système jusque-CRISPR/Cas9 est décrit, y compris les différentes étapes impliquées dans l’Assemblée, purification, concentration et titrage de IDLVs, ainsi que des stratégies pour valider l’efficacité de gène-édition de ces vecteurs. Ce protocole est facilement évolutif pour répondre aux besoins des différents enquêteurs et est conçu pour générer avec succès les vecteurs LV avec des titres dans l’ordre de 1 x 1010 unités (TU) de transduction / mL. Les vecteurs des vicitimes du présent protocole peuvent être utilisés pour infecter efficacement plusieurs types de cellules différentes, y compris de difficile-à-transduce les cellules souches embryonnaires, les cellules hématopoïétiques (les cellules T et les macrophages) et mis en culture et en vivo– neurones injectés. En outre, le protocole est également bien adapté pour la production de vecteurs LENTIVIRAUX intégrase-compétente en quantités similaires.
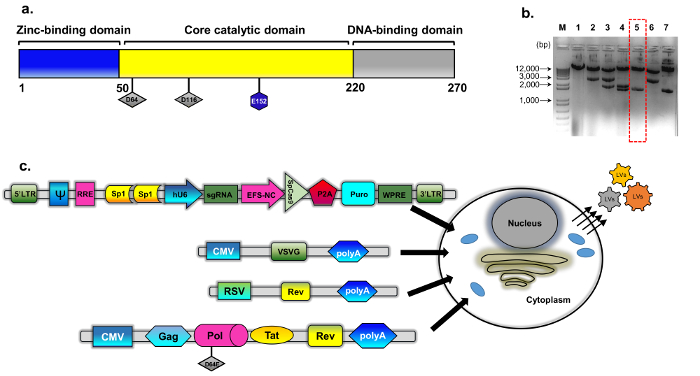
Figure 1 : emballage jusque. (a) schéma du type sauvage intégrase protéine (b) le plasmide modifié a été dérivé de psPAX2 (voir méthodologie, construction plasmidique pour plus de détails). Image de gel d’agarose représentant des clones sélectionnés pour les clones de l’intégrase muté. Des échantillons d’ADN préparés à l’aide d’un mini-kit isolement ADN plasmidique standard ont été analysés par digestion avec EcoRV et SphI. Le clone correctement digérés (numéro 5, boîte rouge en pointillés) a été davantage vérifié par séquençage direct (Sanger) pour la substitution de D64E en INT. La cassette de l’intégrase déficients en emballage s’appelait pBK43. (c) schéma du protocole de transfection transitoire utilisées pour générer des vecteurs jusque-CRISPR/Cas9, montrant des cellules 293 t transfectées avec VSV-G, emballage et cassettes transgène (Sp1-CRISPR/Cas9 tout-en-un plasmide). Les particules virales qui bourgeonnent en dehors de la membrane cellulaire contiennent l’ARN pleine longueur du vecteur (exprimé de la cassette du transgène). La deuxième génération du système d’emballage jusqu’a été utilisée, qui comprend les protéines régulatrices Tat et expression Rev Rev. est complétée d’une cassette séparée (RSV-REV-plasmide). ABBREV: virus de répéter, VSV-G, la stomatite vésiculeuse LTR-Long-terminal G-protéine, promoteur du CMVp-cytomégalovirus ; Promoteur de sarcome de Rous de virus (VRS) ; RRE-(élément de réponse de Rev). Autres éléments régulateurs sur la cassette d’expression comprennent des sites de liaison du Sp1, élément Rev Response (RRE), Woodchuck Hepatitis Virus post-transcriptionnel réglementation élément (WPRE), un promoteur de 1α facteur core-élongation (EFS-NC), l’emballage de vecteur élément ψ (lb/po2), promoteur de cytomégalovirus (hCMV) humain et humain promoteur U6. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
