सटीक जीन संपादन प्रमुख जैव चिकित्सा अग्रिमों कि उपंयास रणनीतियों के विकास के आनुवंशिक रोगों से निपटने के लिए शामिल की आधारशिला रूपों । जीन संपादन प्रौद्योगिकियों के सबसे आगे में सीlustered आरegularly के उपयोग पर निर्भर विधि है-interspaced sप्र॰ palindromic repeats (CRISPR)/Cas9 प्रणाली है कि शुरू में पहचान की गई थी वायरल आनुवंशिक सामग्री के आक्रमण के खिलाफ बैक्टीरियल प्रतिरक्षा के एक घटक के रूप में (संदर्भ1,2) में समीक्षा की । इस तरह के जस्ता के रूप में अंय जीन संपादन उपकरण, पर CRISPR/Cas9 प्रणाली का एक प्रमुख लाभ फिंगर nucleases (ZFNs) और प्रतिलेखन उत्प्रेरक-जैसे प्रभाव nucleases (TALENs) (संदर्भ में समीक्षित3), प्लाज्मिड डिजाइन के सापेक्ष सादगी है और CRISPR घटकों के निर्माण-एक विशेषता है कि एक बहुत व्यापक अनुसंधान समुदाय के लिए कुछ विशेष प्रयोगशालाओं से जीन संपादन के विस्तार संचालित है । इसके अतिरिक्त, CRISPR/Cas9 प्रोग्रामिंग की सादगी और मल्टीप्लेक्सीय लक्ष्य मांयता के लिए इसकी क्षमता को और अधिक प्रभावी और आसान करने के लिए उपयोग प्रौद्योगिकी के रूप में अपनी लोकप्रियता ईंधन है । विभिंन शोधकर्ताओं के लिए उपलब्ध तरीकों के अलावा ऐसी जीन कोशिकाओं को संपादन घटक देने के लिए, वायरल वैक्टर अब तक का सबसे लोकप्रिय और कुशल प्रणाली रहते हैं ।
Lentiviral वैक्टर (LVs) विविध अनुप्रयोगों4,5,6,7के लिए vivo में CRISPR/Cas9 प्रणाली के घटकों को वितरित करने के लिए पसंद के वाहन के रूप में उभरा है । कई प्रमुख विशेषताएं LVs इस प्रक्रिया के लिए एक लोकप्रिय विकल्प बनाने के लिए दोनों विभाजन और गैर विभाजित कोशिकाओं को संक्रमित करने की क्षमता सहित, कम immunogenicity, और ंयूनतम सेलुलर विषाक्तता (संदर्भ में समीक्षा की8) । नतीजतन, एल. वी. मध्यस्थता जीन चिकित्सा संक्रामक रोगों के उपचार में कार्यरत किया गया है, जैसे एचआईवी-1, एचबीवी, और एचएसवी-1, और साथ ही मानव वंशानुगत रोगों अंतर्निहित दोषों के सुधार में, जैसे सिस्टिक फाइब्रोसिस और नव संवहनी धब्बेदार अध… 4 , 5 , 7 , 9 , 10 , 11. इसके अलावा, LVs प्रभावी रूप से अलग जीनोमिक loci पर मल्टीप्लेक्स जीन संपादन प्रदर्शन के लिए एक एकल वेक्टर प्रणाली12का उपयोग कर संशोधित किया गया है ।
हालांकि, LVs के अंतर्निहित संपत्ति मेजबान जीनोम में एकीकृत करने के लिए mutagenic जा सकता है और अक्सर transgene प्रसव के वाहनों के रूप में उनकी उपयोगिता बाधाएं, नैदानिक सेटिंग्स में विशेष रूप से । इसके अलावा, के बाद से छुरा एकीकृत LVs बनाए रखने के उच्च स्तर पर अपने transgenes एक्सप्रेस, इस प्रणाली के बीमार जैसे CRISPR/Cas9 के रूप में जीन संपादन घटकों के वितरण के लिए अनुकूल है; Cas9 के एक्सप्रेस-गाइड आरएनए (gRNA), और ऐसे ZFNs के रूप में इसी तरह के प्रोटीन, बंद लक्ष्य प्रभाव है, जो अवांछनीय उत्परिवर्तनों13,14,15,16 शामिल के ऊंचा स्तर के साथ जुड़े रहे हैं , 17 और संभावित cytotoxicity18में वृद्धि कर सकते हैं । इसलिए, ंयूनतम बंद लक्ष्य प्रभाव के साथ सटीक जीन संपादन को प्राप्त करने के लिए, यह प्रणाली है कि जीन संपादन घटकों के क्षणिक अभिव्यक्ति के लिए अनुमति डिजाइन करने के लिए आवश्यक है ।
हाल के वर्षों में, कई प्रसव प्लेटफार्मों के लिए विकसित किया गया है क्षणिक एक्सप्रेस CRISPR/Cas9 कोशिकाओं में16,19,20,21 (संदर्भ में समीक्षा की22) । इन तरीकों है कि सीधे कोशिकाओं में उपयुक्त गाइड RNAs के साथ शुद्ध Cas9 शुरू करने पर भरोसा शामिल है, जो दिखाया गया था और अधिक प्रभावी जीन प्लाज्मिड की तुलना में संपादन-अभिकर्मक की मध्यस्थता के लिए16. अध्ययनों से पता चला है कि ribonucleoprotein (RNP) गाइड आरएनए से मिलकर परिसरों/Cas9 कणों तेजी से अपने लक्ष्य पर डीएनए दरार मध्यस्थता के बाद बदल रहे हैं, सुझाव है कि इन घटकों की अल्पकालिक अभिव्यक्ति को प्राप्त करने के लिए पर्याप्त है मजबूत जीन संपादन16. adeno-जुड़े वायरल वैक्टर (AAVs) के रूप में गर्भ धारण, गैर एकीकृत वायरल वेक्टर प्लेटफार्मों कोशिकाओं को जीन संपादन मशीनरी देने के लिए एक व्यवहार्य विकल्प प्रदान कर सकते हैं । दुर्भाग्यवश, AAV capsids LVs (< 5kb) की तुलना में काफी कम पैकेजिंग क्षमता रखते हैं, जो किसी एकल सदिश के भीतर बहु-घटक CRISPR टूलकिट को पैकेज करने की क्षमता को गंभीर रूप से बाधित करती है (संदर्भ8में) । यह ध्यान देने योग्य है कि यौगिकों कि हिस्टोन deacetylases (जैसे, सोडियम butyrate23) को बाधित या कोशिका चक्र में बाधा के अलावा (जैसे, कैफीन24) lentiviral titers बढ़ाने के लिए दिखाया गया है । हाल ही में प्रगति के बावजूद, क्षणिक अभिव्यक्ति प्रणालियों अब तक विकसित अब भी कम उत्पादन क्षमता है, जो कम वायरल titers के लिए सीसा, और के माध्यम से उत्पंन वायरस की कम transduction क्षमता के रूप में कई कमियों, द्वारा बाधित कर रहे है इस तरह के दृष्टिकोण25।
Integrase की कमी lentiviral वैक्टर (IDLVs) जीन प्रसव के वाहनों के विकास में एक प्रमुख उंनति का प्रतिनिधित्व करते हैं, के रूप में वे कोशिकाओं में AAV की तरह episomal रखरखाव के अतिरिक्त लाभ के साथ LVs की पैकेजिंग क्षमता गठबंधन । इन सुविधाओं IDLVs काफी हद तक संभावित genotoxic तत्वों और एकीकरण-मध्यस्थता mutagenicity की तुलना लगातार व्यक्त वैक्टर घालमेल के साथ जुड़े प्रमुख मुद्दों को दरकिनार मदद करते हैं । यह पहले से प्रदर्शित किया गया है कि IDLVs सफलतापूर्वक episomal जीन अभिव्यक्ति26,27बढ़ाने के लिए संशोधित किया जा सकता है । साथ IDLV के संबंध में मध्यस्थता CRISPR/Cas9 वितरण, कम उत्पादन titers और episome-integrase-कुशल lentiviral प्रणालियों के सापेक्ष वहन जीनोम के कम अभिव्यक्ति के जीनोम-संपादन देने के लिए बोना के रूप में अपने उपयोगिता को सीमित उपकरण ट्रांसजेनिक construction. हम हाल ही में प्रदर्शित किया है कि दोनों transgene अभिव्यक्ति और वायरल IDLV उत्पादन के साथ जुड़े titers वायरल अभिव्यक्ति की कैसेट28के भीतर प्रतिलेखन कारक Sp1 के लिए बाध्यकारी साइटों के शामिल किए जाने से काफी बढ़ा रहे हैं । संशोधित IDLVs मजबूती से समर्थित CRISPR-मध्यस्थता जीन संपादन दोनों इन विट्रो में (HEK-293T कोशिकाओं में) और vivo में (पोस्ट-mitotic मस्तिष्क न्यूरॉन्स में), जबकि संगत ICLV की तुलना में कम लक्ष्य उत्परिवर्तन उत्प्रेरण-मध्यस्थता सिस्टम28। कुल मिलाकर, हम एक उपंयास, कॉंपैक्ट, सभी में एक CRISPR toolkit एक IDLV मंच पर किया और बढ़ाया जीन संपादन के लिए इस तरह के एक प्रसव के वाहन का उपयोग करने के विभिंन लाभों को रेखांकित विकसित की है ।
यहां, IDLV-CRISPR/Cas9 प्रणाली के उत्पादन प्रोटोकॉल विभिंन विधानसभा में शामिल कदम, शुद्धि, एकाग्रता, और IDLVs के अनुमापन, साथ ही साथ रणनीतियों इन वैक्टर के जीन संपादन प्रभावकारिता को मांय करने सहित वर्णित है । इस प्रोटोकॉल को आसानी से विभिंन जांचकर्ताओं की जरूरतों को पूरा करने के लिए स्केलेबल है और सफलतापूर्वक 1 x 1010 transducing इकाइयों (TU) की श्रेणी में titers के साथ LV वैक्टर उत्पंन करने के लिए डिज़ाइन किया गया है/mL. इस प्रोटोकॉल के माध्यम से उत्पंन वैक्टर कुशलतापूर्वक कई विभिंन प्रकार के सेल को संक्रमित करने के लिए उपयोग किया जा सकता है, मुश्किल से transduce भ्रूण स्टेम सेल, टेम कोशिकाओं (टी कोशिकाओं और मैक्रोफेज), और संस्कृति और vivo मेंशामिल- इंजेक्ट न्यूरॉन्स । इसके अलावा, प्रोटोकॉल समान रूप से अच्छी तरह से integrase के उत्पादन के लिए अनुकूल है-सक्षम lentiviral वैक्टर इसी तरह की मात्रा में ।
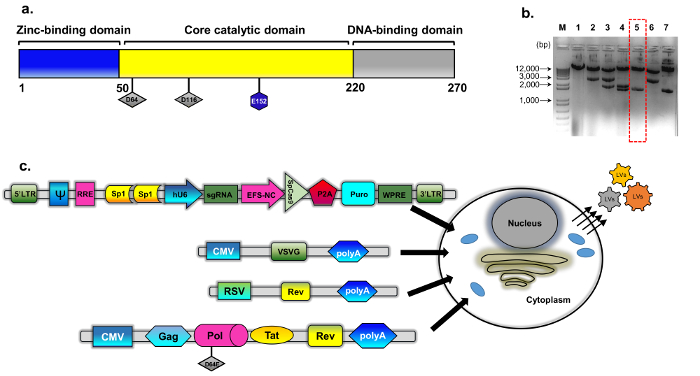
चित्र 1: IDLV पैकेजिंग (क) जंगली प्रकार integrase प्रोटीन की योजनाबद्ध (ख) संशोधित प्लाज्मिड psPAX2 से व्युत्पंन (तरीकों देखें, विवरण के लिए प्लाज्मिड निर्माण) था । प्रतिनिधि agarose जेल रूपांतरित integrase क्लोन के लिए स्क्रीन क्लोन की छवि । डीएनए नमूने एक मानक प्लाज्मिड डीएनए अलगाव मिनी किट का उपयोग कर तैयार EcoRV और SphI के साथ पाचन द्वारा विश्लेषण किया गया । सही ढंग से पचा क्लोन (संख्या 5, लाल बॉक्स धराशायी) आगे INTमें D64E प्रतिस्थापन के लिए प्रत्यक्ष (सैंज) अनुक्रमण द्वारा सत्यापित किया गया था । integrase की कमी वाली पैकेजिंग कैसेट का नाम pBK43 था. (ग) IDLV-CRISPR/Cas9 वैक्टर उत्पन्न करने के लिए कार्यरत क्षणिक अभिकर्मक प्रोटोकॉल की योजनाबद्ध, 293T के साथ transfected कोशिकाओं को दिखा-जी, पैकेजिंग, और VSV कैसेट्स (Sp1-transgene/CRISPR ऑल-इन-वन Cas9). वायरल कणों कि कोशिका झिल्ली से बाहर कली सदिश की पूरी लंबाई आरएनए होते है (transgene कैसेट से व्यक्त) । IDLV-पैकेजिंग प्रणाली की दूसरी पीढ़ी है, जो विनियामक प्रोटीन जैसे और rev. rev अभिव्यक्ति आगे एक अलग कैसेट (RSV-rev-प्लाज्मिड) से पूरक है शामिल किया गया था । Abbrev: लीटर लंबी टर्मिनल दोहराने, VSV-जी, vesicular stomatitis वायरस जी प्रोटीन, pCMV-cytomegalovirus प्रवर्तक; रोस सार्कोमा वायरस (RSV) प्रवर्तक; RRE-(Rev प्रतिक्रिया तत्व) । अभिव्यक्ति कैसेट पर अंय विनियामक तत्वों Sp1-बाध्यकारी साइटों, Rev प्रतिक्रिया तत्व (RRE), Woodchuck हेपेटाइटिस वायरस Posttranscriptional विनियामक तत्व (WPRE), एक कोर-बढ़ाव कारक 1α प्रमोटर (EFS-नेकां), वेक्टर पैकेजिंग शामिल तत् ψ (psi), ह्यूमन Cytomegalovirus (hCMV) प्रवर्तक, और मानव U6 प्रवर्तक । कृपया यहां क्लिक करें इस आंकड़े का एक बड़ा संस्करण को देखने के लिए ।
