Presis genet redigering utgjør hjørnesteinen i store biomedisinsk fremskritt som involverer utviklingen av romanen strategier for å takle genetiske sykdommer. I forkant av gen-redigering teknologiene er metoden stole på bruk av den clustered regularly –jegnterspaced short palindromic repeats (CRISPR) / Cas9 system som ble identifisert som en del av bakteriell immunitet mot invasjonen av viral genetiske materiale (omtalt i referanser1,2). En stor fordel av CRISPR/Cas9 over andre gen-redigeringsverktøy, som sink-finger nucleases (ZFNs) og transkripsjon aktivator som effektor nucleases (TALENs) (omtalt i referanse3), er den relative enkelheten av plasmider design og bygging av CRISPR komponenter, en funksjon som har drevet utvidelse av gen-redigering av noen få spesialiserte laboratories til en mye bredere forskningsmiljø. I tillegg har enkelheten til CRISPR/Cas9 programmering og dens kapasitet for multiplex målet anerkjennelse ytterligere fueled sin popularitet som en kostnadseffektiv og lett-å-bruke teknologi. Blant de ulike metodene tilgjengelig for forskere å levere slike gen-redigering komponenter til celler, fortsatt viral vektorer langt den mest populære og effektive systemet.
Lentiviral vektorer (LVs) har dukket opp som den ønsker å levere komponentene i CRISPR/Cas9 systemet i vivo for diverse programmer4,5,6,7. Flere viktige funksjoner gjør LVs et populært valg for denne prosessen inkluderer deres evne til å infisere både dele og ikke dele celler, lav immunogenisitet og minimal mobilnettet toksisitet (omtalt i referanse8). Som et resultat, har LV-mediert genterapi vært ansatt i behandling av infeksjonssykdommer, som HIV-1 og HBV HSV-1, og i korrigering av feil underliggende menneskelige arvelige sykdommer, som cystisk fibrose og neo-vaskulær macular degenerasjon 4 , 5 , 7 , 9 , 10 , 11. videre, LVs har endret effektivt for å utføre multiplex genet redigering på tydelige genomisk loci bruker en enkelt vektor systemet12.
Men den iboende egenskapen av LVs å integrere i vert genomet kan mutagene og ofte handicap deres nytte som transgene levering biler, spesielt i klinisk innstillinger. Videre siden stabilt integrert LVs uttrykke deres effekter av transgener på bærekraftig høye nivåer, er dette systemet dårlig tilpasset for levering av gen-redigering komponenter som CRISPR/Cas9; overuttrykte Cas9-guide RNA (gRNA) og lignende proteiner som ZFNs, er forbundet med forhøyede nivåer av off-målet effekter, blant annet uønsket mutasjoner13,14,15,16 , 17 og kan potensielt øke cytotoksisitet18. Derfor, for å oppnå presis gen-redigering med minimal off-målet effekter, er det viktig å design systemer som tillater for forbigående uttrykket av genet redigeringskomponenter.
De siste årene, en rekke levering plattformer er utviklet for å uttrykke transiently CRISPR/Cas9 i cellene16,19,20,21 (omtalt i referanse22). Disse omfatter metoder som bruker direkte innføre renset Cas9 sammen med riktig veiledning RNAs i celler, som viste seg å være mer effektiv på målrettet gen-redigering i forhold til plasmider-mediert transfection16. Studier har vist at ribonucleoprotein (RNP) komplekser bestående av guide RNA/Cas9 partikler er raskt snudd etter formidling DNA cleavage på sine mål, antyder at kortsiktige uttrykket av disse komponentene er tilstrekkelig for å oppnå robust genet redigering16. Tenkes, ikke integrert viral vektor plattformer som adeno-assosiert virus vektorer (AAVs) kan gi et levedyktig alternativ å levere gen-redigering maskiner til cellene. Dessverre AAV capsids har betydelig lavere emballasje kapasiteter enn LVs (< 5kb), som sterkt hindrer deres evne til å pakke multi-komponent CRISPR verktøysettet innenfor en enkelt vektor (omtalt i referanse8). Det er verdt å merke seg at tillegg av forbindelser som hemme histone deacetylases (f.eks, natrium butyrate23) eller hindre cellen syklus (f.eks, koffein24) har vist seg å øke lentiviral titers. Til tross for framskritt, er forbigående uttrykk systemer utviklet så langt fortsatt hindret av flere mangler, for eksempel lavere effektivitet, som fører til redusert viral titers, og lav signaltransduksjon effektiviteten av virus generert gjennom slike tilnærminger25.
Integrase dårlig lentiviral vektorer (IDLVs) representerer et stort fremskritt i utviklingen av genet levering biler, som de kombinerer emballasje evnen til LVs med fordelen av AAV-lignende episomal vedlikehold i celler. Disse funksjonene hjelper IDLVs i stor grad omgå de store problemene forbundet med å integrere vektorer, vis-à-vis kontinuerlig overuttrykte potensielt gentoksisk elementer og integrasjon-mediert genetisk virkning. Det ble tidligere vist at IDLVs kan endres med hell for å forbedre episomal gene expression26,27. Med hensyn til IDLV-mediert CRISPR/Cas9 levering begrenser lav produksjon titers og lavere uttrykk for episome-borne genomer i forhold til integrase-dyktige lentiviral systemer deres nytte som bona fide verktøy for å levere genomet-redigering transgene konstruksjoner. Vi har nylig vist at både transgene uttrykk og viral titers knyttet IDLV produksjon er betydelig forbedret ved inkludering av bindende områder for transkripsjon factor Sp1 i viral uttrykk kassett28. De endrede IDLVs støttet robust CRISPR-mediert genet redigering både i vitro (i HEK-293T celler) og i vivo (på etter mitotisk hjernen nevroner), mens inducing minimal off-målet mutasjoner sammenlignet med tilsvarende ICLV-mediert systemer28. Samlet, vi utviklet en roman, kompakt, alle-inne-ettall CRISPR toolkit gjennomført på en IDLV plattform og skissert ulike fordelene med å bruke slike en levering kjøretøy for forbedret genet redigering.
Her er produksjon protokollen av IDLV-CRISPR/Cas9 beskrevet, inkludert de ulike trinnene involvert i samlingen, renselse, konsentrasjon og titrering av IDLVs, samt strategier for å validere gen-redigering effekten av disse vektorer. Denne protokollen er lett skalerbare for å møte behovene til ulike etterforskere og er utformet for å kunne generere LV vektorer med titers i området fra 1 x 1010 transducing enheter (TU) / mL. Vektorer generert gjennom denne protokollen kan brukes til å effektivt infisere flere ulike celletyper, inkludert vanskelig å transduce embryonic stilk celler, blodkreft cellene (T-celler og makrofager) og kultivert og i vivo– injisert neurons. Videre er protokollen like godt egnet for produksjon av integrase-kompetent lentiviral vektorer i lignende mengder.
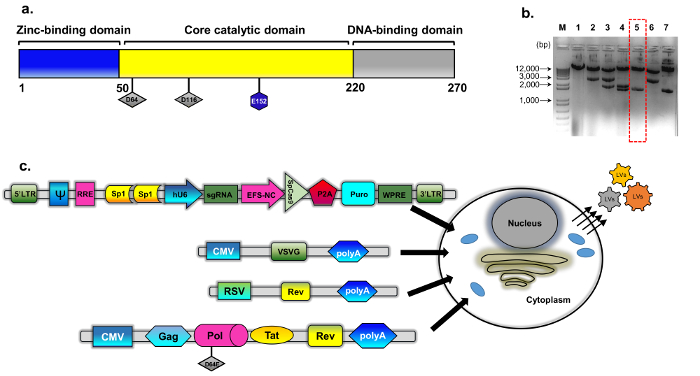
Figur 1: IDLV emballering. (a) skjematisk av wild type integrase protein (b) den endrede plasmider ble avledet fra psPAX2 (se metoder, plasmider konstruksjon for detaljer). Representant agarose gel bilde av kloner vist for muterte integrase kloner. DNA-prøver tilberedt med en standard plasmider DNA isolasjon mini kit ble analysert av fordøyelsen med EcoRV og SphI. Riktig fordøyd klone (nummer 5, stiplet rød boks) ble ytterligere bekreftet ved direkte (Sanger) sekvenser for D64E substitusjon i INT. Integrase-mangelfull emballasje kassetten ble kalt pBK43. (c) skjematisk av forbigående transfection protokollen ansatt til å generere IDLV-CRISPR/Cas9 vektorer, viser transfekterte 293T celler med VSV-G, emballasje og transgene kassetter (Sp1-CRISPR/Cas9 alt-i-ett plasmider). Virus partikler som bud ut fra cellemembranen inneholder den fulle RNA av vektoren (uttrykt fra transgene kassetten). Den andre generasjonen av IDLV-emballasje systemet ble brukt, som inkluderer regulatoriske proteiner Tat og rev Rev uttrykk er ytterligere supplert fra en egen kassett (RSV-REV-plasmider). ABBREV: LTR-lang-terminal gjenta, VSV-G, vesicular stomatitt virus G-protein, pCMV-cytomegalovirus promoter; Rous sarkomspesialitet virus (RSV) selskapet; RRE-(Rev svar Element). Andre regulerende elementer på uttrykk kassetten inkluderer Sp1 bindende områder, Rev Response element (RRE), hakkespett hepatitt Virus Posttranscriptional regulatorisk Element (WPRE), en kjerne-forlengelse faktor 1α promoter (EFS-NC), vektor emballasjen elementet ψ (psi), menneskelige Cytomegalovirus (hCMV) arrangøren og menneskelige U6 promoter. Klikk her for å se en større versjon av dette tallet.
