A segmentação metamérica de organismos é amplamente utilizada na natureza. Estruturas repetidas são essenciais para a funcionalidade de órgãos laterais como vértebras, músculos, nervos, vasos, membros ou folhas em um plano corporal1. Como resultado de tais restrições fisiológicas e geométricas da simetria axial, a maioria da phyla de Bilateria- como annelids, artrópodes e cordatas-exposição segmentação de seus tecidos embrionários (por exemplo, ectoderme, mesoderme) antero-posteriormente.
Embriões vertebrados segmentam sequencialmente seu mesoderme paraxial ao longo do eixo principal do corpo em somitas com intervalos específicos de espécies, contagens e distribuições de tamanho. Apesar de tal robustez entre embriões individuais dentro de uma espécie, a segmentação de somite é versátil entre espécies de vertebrados. A segmentação acontece em um vasto regime de intervalos de tempo (de 25 minutos em zebrafish a 5 h em humanos), tamanhos (de ~20 μm em somitas de cauda de zebrafish a ~200 μm em somitas tronco de camundongos) e conta (de 32 em zebrafish a ~300 em cobras de milho)2. O mais interessante é que os embriões de peixe podem desenvolver-se em uma ampla gama de temperaturas (de ~20,5 °C até 34 °C para zebrafish) mantendo seus somitas intactos com distribuições de tamanho adequado, compensando tanto os intervalos de segmentação quanto as velocidades de alongamento axial. Além dessas características interessantes, o zebrafish permanece como um organismo modelo útil para estudar a segmentação em vertebrados devido ao desenvolvimento externo, síncrono e transparente de uma plenitude de embriões irmãos, bem como suas ferramentas genéticas acessíveis. Adversamente a partir de uma perspectiva de microscopia, os embriões teleost desenvolvem-se em uma gema esférica volumosa, esticando e arredondando o tecido gastrulating ao seu redor(Figura 1A). Neste artigo, apresentamos uma cultura de explante de tecido 3D achatada para caudas de zebrafish. Este sistema de explant contorna as restrições esféricas da massa de gema, permitindo o acesso a imagens vivas de alta resolução de embriões de peixes para padronização de somite.
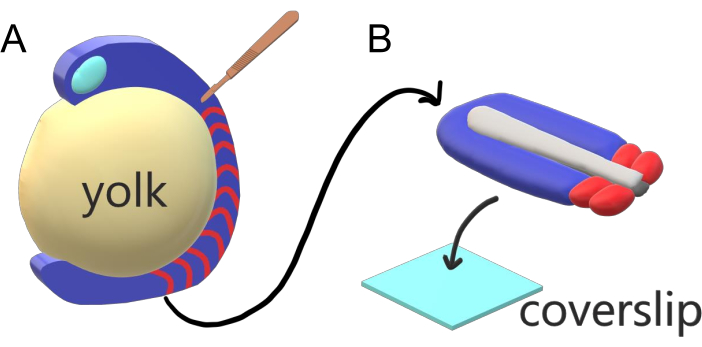
Figura 1: Sistema de Explantação de Câmara de Slides para Embriões de Zebrafish. (A) Os embriões de zebrafish têm vantagens para imagens vivas, como a transparência do tecido embrionário gastrulating (azul), mas o tecido se forma em torno de uma massa de gema esférica volumosa (amarela) que previne imagens quase objetivas e de alta resolução em embriões intactos. Explantas de cauda podem ser dissecadas começando com uma faca microcirúrgica (marrom) cortada do tecido anterior de somites (vermelho) e continuando na borda com a gema posteriormente. (B) Explantas traseiras dissecadas podem ser colocadas em um deslizamento (azul claro) dorsoventrally; mantendo tecido neural (cinza claro) em cima e notochord (cinza escuro) na parte inferior. Clique aqui para ver uma versão maior desta figura.
