| General Materials |
|
|
|
| 500 mL Bottletop Filter .22 µm PES Sterilizing, Low Protein Binding, w/45mm Neck |
Fisher |
CLS431154 |
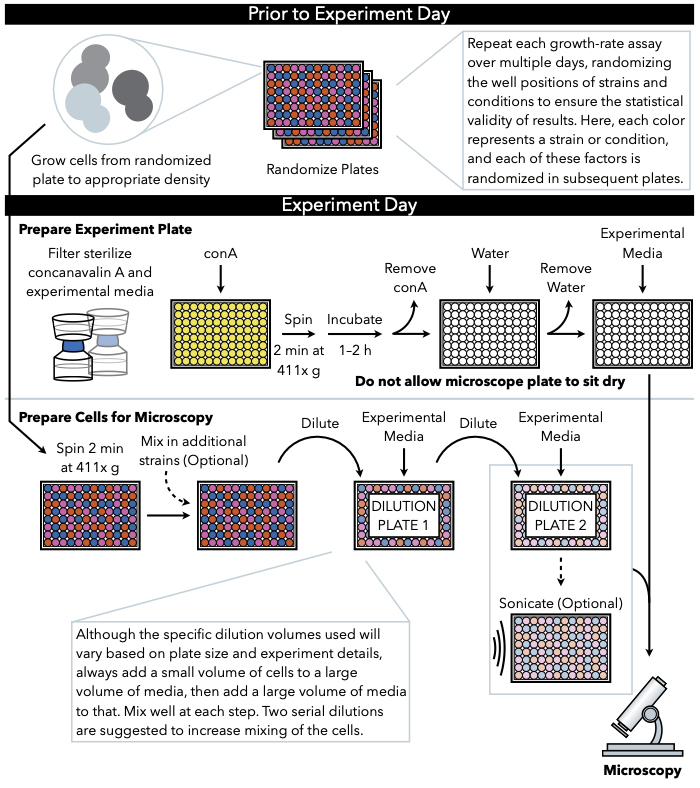
used to filter the media |
| BD Falcon*Tissue Culture Plates, microtest u-bottom |
Fisher |
08-772-54 |
96-well culture tubes used to freeze cells, pre-grow cells, and dilutions |
| BD Syringes without Needle, 50 mL |
Fisher |
13-689-8 |
Used to filter the Concanavalin A |
| Costar Sterile Disposable Reagent Reservoirs |
Fisher |
07-200-127 |
reagent reservoirs used to pipette solutions with multichannel pipette |
| Costar Thermowell Aluminum Sealing Tape |
Fisher |
07-200-684 |
96-well plate seal for pre-growth and freezing |
| lint and static free Kimwipes |
Fisher |
06-666A |
lint and static free wipes to keep microscope plate bottom free of debris and scratches |
| Nalgene Syringe Filters |
ThermoFisher Scientific |
199-2020 |
0.2 μm pore size, 25 mm diameter; used to filter concanavalin A solution |
| Media Components |
|
|
|
| Minimal chemically defined media (MD; 2% glucose) |
|
|
alternative microscopy media used for yeast pre-growth and growth during microscopy |
| Synthetic Complete Media (SC; 2% glucose) |
|
|
microscopy media used for yeast pre-growth and growth during microscopy |
| Yeast extract-peptone-dextrose (YEPD; 2% glucose) medium |
|
|
cell growth prior to freezing down randomized plates |
| Microscopy Materials |
|
|
|
| Breathe-Easy sealing membrane |
Millipore Sigma |
Z380059-1PAK |
breathable membranes used to seal plate during microscopy experiment. At this stage breathable membranes are reccomended because they prevent condensation in the wells and allow for better microscopy images |
| Brooks 96-well flat clear glass bottom microscope plate |
Dot Scientific |
MGB096-1-2-LG-L |
microscope plate |
| Concanavalin A from canavalia ensiformis (Jack Bean), lyophilized powder |
Millipore Sigma |
45-C2010-1G |
Make 5x concanavalin A solution and freeze 5ml of 5x concanavalin A in 50 mL conical tubes at -80 °C |
| Strains Used |
|
|
|
| MAH.5, MAH.96, MAH.52, MAH.66, MAH.11, MAH.58, MAH.135, MAH.15, MAH.44, MAH.132 |
|
|
Haploid mutation accumulation strains in a laboratory background, described in Hall and Joseph 2010 |
| EP026.2A-2C |
|
|
Progeny of the ancestral Hall and Joseph 2010 mutation accumulation strain, transformed with YFR054cΔ::Scw11P::GFP |
| Equipment |
|
|
|
| Misonix Sonicator S-4000 with 96-pin attachment |
|
|
Sonicator https://www.labx.com/item/misonix-inc-s-4000-sonicator/4771281 |
| Nikon Eclipse Ti-E with Perfect Focus System |
|
|
Inverted microscope with automated stage and autofocus system |