Vekst fenotyper bidrar kritisk til gjær fitness. Naturlig utvalg kan effektivt skille mellom avstamninger med vekstrater forskjellig fra det motsatte av den effektive befolkningsstørrelsen, som kan overstige 108 personer1. Videre er variasjon av vekstrater blant individer i en befolkning en evolusjonært relevant parameter, da den kan tjene som grunnlag for overlevelsesstrategier sominnsatssikring 2,3,4,5,6. Derfor er analyser som tillater svært nøyaktige målinger av vekstfenotyper og deres fordelinger avgjørende for studiet av mikroorganismer. Mikrokolonivekstanalysen som er beskrevet her, kan generere individuelle vekstratemålinger for ~ 105 mikrokolonier per eksperiment. Denne analysen gir derfor en kraftig protokoll for å studere gjær evolusjonær genetikk og genomikk. Det egner seg spesielt godt til å teste hvordan variasjon i populasjoner av genetisk identiske enkeltceller genereres, vedlikeholdes og bidrar til befolkningsform7,8,9,10.
Metoden som er beskrevet her (Figur 1), bruker periodisk fangede, lavforstørrelsesbilder av celler som vokser i flytende medier på en 96- eller 384-brønns glassbunnplate for å spore vekst i mikrokolonier. Cellene holder seg til lectin concanavalin A, som dekker bunnen av mikroskopplaten, og danner todimensjonale kolonier. Fordi mikrokoloniene vokser i en monolayer, er mikrokoloniområdet svært korrelert med celle nummer7. Derfor kan nøyaktige estimater av mikrokoloni vekstrate og oppholdstid genereres med tilpasset bildeanalyseprogramvare som sporer endringshastigheten for området for hvert mikrokoloni. Videre kan det eksperimentelle oppsettet overvåke overflod og til og med subcellulære lokaliseringer av fluorescerende merkede proteiner uttrykt i disse mikrokoloniene. Nedstrøms behandling av data fra denne mikrokoloni vekstanalysen kan oppnås ved tilpasset analyse eller av eksisterende bildeanalyseprogramvare, for eksempel Processing Images Easily (PIE)11, en algoritme for robust koloniområdegjenkjenning og vekstanalyse med høy gjennomstrømning fra lavforstørrelse, brightfield-bilder, som er tilgjengelig via GitHub12.
Fordi vekstrateestimater avledet fra mikrokoloni-vekstanalysen genereres fra et stort antall enkeltkolonimålinger, er de ekstremt nøyaktige, med standardfeil flere størrelsesordener mindre enn estimatene selv for et rimelig stort eksperiment. Derfor er analysens kraft til å oppdage vekstratforskjeller mellom forskjellige genotyper, behandlinger eller miljøforhold høy. Multiwell-plateformatet gjør at mange forskjellige miljø- og genotypekombinasjoner kan sammenlignes i et enkelt eksperiment. Hvis stammer uttrykker forskjellige fluorescerende markører, kan de blandes i samme brønn og preges av påfølgende bildeanalyse, noe som kan øke kraften ytterligere ved å tillate brønn-for-brønn data normalisering.
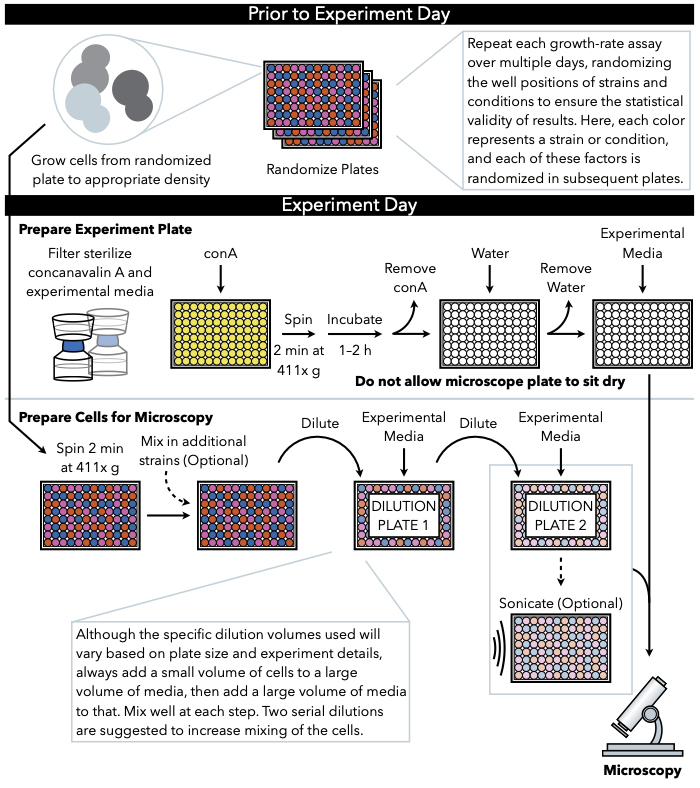
Figur 1: Skjematisk representasjon av protokollen. Denne protokollen følger to hovedtrinn, som er utarbeidelsen av eksperimentell plate og forberedelsen av cellene til bilde. Randomisering av plater og vekst av celler bør utføres før og før eksperimentdagen. Gjentatt blanding av celler på hvert trinn under fortynning er viktig i trinnene til plating, og derfor anbefales det å forberede den eksperimentelle platen først, slik at den er klar til plating umiddelbart etter ferdigstillelse av cellefortynning. Klikk her for å se en større versjon av denne figuren.
