Strongyloides stercoralis wurde lange Zeit als wichtiger menschlicher Krankheitserreger im Vergleich zu den weiter verbreiteten Hakenwürmern und dem Spulwurm Ascaris lumbricoides1 übersehen. Frühere Studien zur Wurmbelastung unterschätzten die Prävalenz von S. stercoralis aufgrund der geringen Sensitivität gängiger Diagnosemethoden für S. stercoralisoft stark 2. In den letzten Jahren haben epidemiologische Studien, die auf verbesserten diagnostischen Instrumenten basieren, geschätzt, dass die tatsächliche Prävalenz von S. stercoralis-Infektionen viel höher ist als zuvor berichtet, etwa 610 Millionen Menschen weltweit2.
Sowohl S. stercoralis als auch andere Strongyloides-Arten, einschließlich des eng verwandten Rattenparasiten und des gemeinsamen Labormodells S. ratti, haben einen ungewöhnlichen Lebenszyklus, der für experimentelle genomische Studien von Vorteil ist, da er sowohl aus parasitären als auch aus freilebenden (Umwelt-) Generationen 3 besteht (Abbildung 1). Insbesondere können sowohl S. stercoralis als auch S. ratti durch eine einzige frei lebende Generation durchlaufen. Die freilebende Generation besteht aus postparasitären Larven, die sich zu frei lebenden erwachsenen Männchen und Weibchen entwickeln; Alle Nachkommen der frei lebenden Erwachsenen entwickeln sich zu infektiösen Larven, die einen Wirt infizieren müssen, um den Lebenszyklus fortzusetzen. Darüber hinaus kann diese ökologische oder frei lebende Generation im Labor experimentell manipuliert werden. Da freilebende Strongyloides-Erwachsene und C. elegans-Erwachsene eine ähnliche Morphologie aufweisen, können Techniken wie die intragonadale Mikroinjektion, die ursprünglich für C. elegans entwickelt wurden, für die Verwendung mit freilebenden erwachsenen Strongyloides 4,5 angepasst werden. Während DNA im Allgemeinen in frei lebende erwachsene Weibchen eingeführt wird, können sowohl Männer als auch Frauen von Strongyloides mikroinjiziert werden6. Somit stehen funktionelle genomische Werkzeuge zur Verfügung, um viele Aspekte der Biologie von Strongyloides zu untersuchen. Anderen parasitären Nematoden fehlt eine frei lebende Generation und sie sind daher nicht so leicht für funktionelle genomische Technikengeeignet 3.
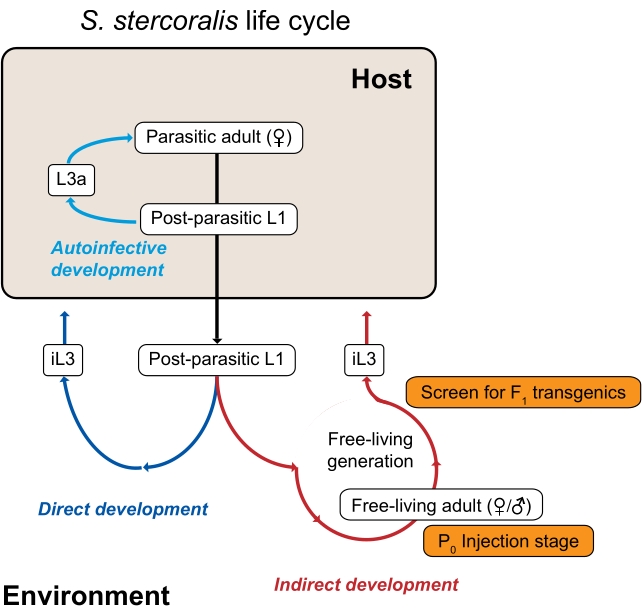
Abbildung 1: Der Lebenszyklus von Strongyloides stercoralis. Die parasitären Weibchen von S. stercoralis bewohnen den Dünndarm ihrer Säugetierwirte (Menschen, nichtmenschliche Primaten, Hunde). Die parasitären Weibchen vermehren sich durch Parthenogenese und legen Eier in den Dünndarm. Die Eier schlüpfen, während sie sich noch im Inneren des Wirts befinden, zu postparasitären Larven, die dann mit Kot in die Umwelt gelangen. Wenn die postparasitären Larven männlich sind, entwickeln sie sich zu frei lebenden erwachsenen Männchen. Wenn die postparasitären Larven weiblich sind, können sie sich entweder zu frei lebenden erwachsenen Weibchen (indirekte Entwicklung) oder zu infektiösen Larven im dritten Stadium (iL3s; direkte Entwicklung) entwickeln. Die frei lebenden Männchen und Weibchen vermehren sich sexuell, um Nachkommen zu schaffen, die gezwungen sind, iL3s zu werden. Unter bestimmten Bedingungen kann S. stercoralis auch einer Autoinfektion unterzogen werden, bei der einige der postparasitären Larven im Darm des Wirts verbleiben, anstatt im Kot in die Umwelt überzugehen. Diese Larven können sich im Inneren des Wirts zu autoinfektiösen Larven (L3a) entwickeln, durch die Darmwand eindringen, durch den Körper wandern und schließlich in den Darm zurückkehren, um reproduktive Erwachsene zu werden. Der Lebenszyklus von S. ratti ist ähnlich, außer dass S. ratti Ratten infiziert und keinen autoinfektiösen Zyklus hat. Die Umweltgenerierung ist der Schlüssel zur Verwendung von Strongyloides-Arten für genetische Studien. Die frei lebenden erwachsenen Weibchen (P0) können mikroinjiziert werden; ihre Nachkommen, die alle zu iL3s werden, sind die potenziellen F 1-Transgenen. Diese Figur wurde von Castelletto et al. modifiziert. 3. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
S. stercoralis teilt viele Aspekte seiner Biologie mit anderen gastrointestinalen menschlich-parasitären Nematoden, einschließlich Wirtsinvasion und Wirtsimmunmodulation. Zum Beispiel infizieren sich auch humanparasitäre Hakenwürmer der Gattungen Necator und Ancylostoma durch Hautpenetration, navigieren ähnlich durch den Körper und leben schließlich als parasitäre Erwachsene imDünndarm 7. Daher verwenden viele gastrointestinale Nematoden wahrscheinlich gängige sensorische Verhaltensweisen und Immunevasionstechniken. Infolgedessen wird das aus Strongyloides gewonnene Wissen die Ergebnisse anderer weniger genetisch beherrschbarer Nematoden ergänzen und zu einem vollständigeren Verständnis dieser komplexen und wichtigen Parasiten führen.
Dieses Mikroinjektionsprotokoll beschreibt die Methode zur Einführung von DNA in Strongyloides freilebende erwachsene Frauen, um transgene und mutierte Nachkommen herzustellen. Die Anforderungen an die Aufrechterhaltung des Stammes, einschließlich des Entwicklungszeitpunkts von erwachsenen Würmern für Mikroinjektionen und der Sammlung transgener Nachkommen, werden beschrieben. Protokolle und eine Demonstration der vollständigen Mikroinjektionstechnik sowie Protokolle für die Kultivierung und das Screening transgener Nachkommen sind enthalten, zusammen mit einer Liste aller notwendigen Geräte und Verbrauchsmaterialien.
