אינטראקציות ביומולקולריות:
חלבונים הם חלקים חיוניים של אורגניזמים ומשתתפים במסלולים מולקולריים רבים כגון חילוף חומרים של תאים, מבנה התא, איתות תאים, תגובות חיסוניות, הידבקות תאים ועוד. בעוד שחלבונים מסוימים מבצעים את תפקידם באופן עצמאי, רוב החלבונים מקיימים אינטראקציה עם חלבונים אחרים באמצעות ממשק מקשר כדי לתאם פעילות ביולוגית תקינה1.
ניתן לסווג אינטראקציות ביומולקולריות בעיקר על סמך המאפיינים המבניים והתפקודיים המובהקים של חלבונים המעורבים2, למשל, בהתבסס על משטחי החלבון, היציבות המורכבת או ההתמדה של אינטראקציות3. זיהוי חלבונים חיוניים ותפקידם באינטראקציות ביומולקולריות הוא חיוני להבנת מנגנונים ביוכימיים ברמה המולקולרית4. נכון לעכשיו, ישנן גישות שונות לזיהוי אינטראקציות אלה5: in vitro6, בסיליקו7, בתאים חיים8, ex vivo9, ו in vivo10 כאשר לכל אחד מהם יש חוזקות וחולשות משלו.
מבחני in vivo מבוצעים תוך שימוש בחיה כולה ככלי ניסיוני11, ומבחניה- ex vivo מבוצעים על תמציות רקמה או על איברים שלמים (למשל, לב, מוח, כבד) בסביבה חיצונית מבוקרת על ידי מתן שינויים מינימליים בתנאים טבעיים. היישום הנפוץ ביותר של מחקרי in vivo ו – ex vivo הוא להעריך את הפרמקוקינטיקה, הפרמקודינמיקה והשפעות הרעילות של חומרים פרמקולוגיים פוטנציאליים לפני ניסויים בבני אדם על ידי הבטחת הבטיחות והיעילות הכוללות שלהם12.
ניתן לזהות אינטראקציות ביומולקולריות גם בתוך תאים חיים. הדמיית תאים חיים מאפשרת לנו לצפות באינטראקציות דינמיות בזמן שהם מבצעים את התגובות של מסלול ביוכימי מסוים13. יתר על כן, טכניקות זיהוי, כגון ביולומינסנציה או העברת אנרגיית תהודה פלואורסצנטית, יכולות לספק מידע על היכן ומתי אינטראקציות אלה מתרחשות בתוך תא14. אף על פי שזיהוי בתאים חיים מציע פרטים חיוניים, מתודולוגיות זיהוי אלה מסתמכות על אופטיקה ותוויות, שייתכן שאינן משקפות את הביולוגיה המקורית; הם גם פחות מבוקרים משיטות במבחנה ודורשים מומחיות מיוחדת כדי לבצע15.
השיטות החישוביות של in silico משמשות בעיקר לסינון בקנה מידה גדול של מולקולות מטרה לפני הניסויים במבחנה . שיטות חיזוי חישוביות, מסדי נתונים מבוססי מחשב, עגינה מולקולרית, יחסי מבנה-פעילות כמותיים וגישות סימולציה אחרות של דינמיקה מולקולרית הם בין המבוססים היטב בכלי סיליקו 16. בהשוואה לטכניקות ניסיוניות מייגעות, הכלים בסיליקו יכולים בקלות לבצע תחזיות עם רגישות גבוהה, אך עם דיוק מופחת בביצועי החיזוי17.
בדיקות במבחנה מבוצעות עם מיקרואורגניזמים או מולקולות ביולוגיות מחוץ להקשר הביולוגי הסטנדרטי שלהם. הצגת אינטראקציות ביומולקולריות באמצעות שיטות במבחנה היא קריטית להבנת תפקודי החלבון והביולוגיה שמאחורי הרשת המורכבת של תפקוד התאים. מתודולוגיית הבדיקה המועדפת נבחרת על פי התכונות הפנימיות של החלבון, הערכים הקינטיים והאופן והעוצמה של האינטראקציות18,19.
האינטראקציה Hsp90/Cdc37:
מסלול המלווה-קינאז, המחבר בין Hsp90 ל-Cdc37, הוא מטרה טיפולית מבטיחה בביולוגיה של הגידול20. Hsp90 ממלא תפקיד מרכזי בבקרת מחזור התא, בהרכבת חלבונים, בהישרדות התאים ובמסלולי איתות. חלבונים המסתמכים על Hsp90 לצורך תפקודם מועברים ל-Hsp90 לצורך קומפלקסציה באמצעות מלווה משותף, כגון Cdc37. קומפלקס Hsp90/Cdc37 שולט בקיפול של רוב החלבון קינאזות ומשמש כמרכז לשלל רשתות איתותתוך-תאיות 21. זוהי מטרה אנטי-סרטנית מבטיחה בשל הביטוי המוגבה שלה בממאירות שונות, כולל לוקמיה מיאלובלסטית חריפה, מיאלומה נפוצה וקרצינומה הפטוצלולרית22,23.
טכניקות נפוצות לזיהוי אינטראקציה ביומולקולרית במבחנה
Co-immunoprecipitation (co-IP) היא טכניקה המסתמכת על ספציפיות אנטיגן-נוגדנים כדי לזהות אינטראקציות רלוונטיות ביולוגית24. החיסרון העיקרי של שיטה זו הוא חוסר היכולת שלה לזהות אינטראקציות בעלות זיקה נמוכה וערכים קינטיים24. שיטות ביופיזיות כגון קלורימטריה איזותרמית של טיטרציה (ITC), תהודת פלסמון פני השטח (SPR), אינטרפרומטריה ביו-שכבתית (BLI) וטכנולוגיית FEB עדיפות לקביעת הערכים הקינטיים.
ITC היא שיטת זיהוי ביו-פיזיקלית המבוססת על קביעת אנרגיית קשירת יחד עם ניתוח תרמודינמיקה מלא לאפיון אינטראקציות ביומולקולריות25. היתרון העיקרי של ITC הוא שהוא אינו דורש כל תיוג או קיבוע של חלבון המטרה. הקשיים העיקריים שבהם נתקל ITC הם הריכוז הגבוה של חלבון המטרה הנדרש לניסוי אחד והקושי לנתח קומפלקסים לא קוולנטיים עקב אנתלפיות קישור קטנות26. גם SPR וגם BLI הן טכניקות ביופיזיות נטולות תוויות המסתמכות על אימוביליזציה של מולקולת המטרה על משטח החיישנים, ולאחר מכן זריקות של האנליט מעל המטרה המשותקת27,28. ב- SPR, שינויים במקדם השבירה במהלך אינטראקציות ביומולקולריות נמדדים27; ב-BLI, ההתאבכות באור המוחזר נרשמת בזמן אמת כשינוי באורך הגל כפונקציה של הזמן28. גם SPR וגם BLI חולקים יתרונות משותפים של הצעת יכולות ספציפיות, רגישות וזיהוי גבוהות29. בשתי השיטות, חלבון המטרה משותק על משטחי ביוסנסור, ולכן, ייתכן שיש אובדן מסוים של הקונפורמציה הטבעית של המטרה, מה שמקשה על הבחנה בין אינטראקציות ספציפיות לעומת לא ספציפיות30. BLI משתמשת בביו-סנסורים חד-פעמיים חד-פעמיים יקרים כדי לשתק את המטרה, ולכן היא טכניקה יקרה31. בהשוואה לכלי זיהוי ביומולקולריים מבוססים אלה, טכנולוגיית FEB מציעה פלטפורמה אמינה ונטולת תוויות על ידי שימוש בריכוזי ננומולארים נמוכים לזיהוי ביומולקולרי בזמן אמת עם אפיון קינטי. טכנולוגיית FEB גם מתגברת על האתגרים המבעבעים העומדים בפני ITC והיא חסכונית יותר בהשוואה ל- SPR או BLI.
ביוסנסורים מבוססי טרנזיסטור בעל אפקט שדה (FET) הוא תחום מתפתח לאיתור אינטראקציות ביומולקולריות על ידי הצעת יישומים ביו-רפואיים מגוונים. במערכת FET, מטרות משותקות לשבבי הביו-סנסור ואינטראקציות מזוהות על ידי שינויים בהולכה32. התכונה הייחודית שיש לקחת בחשבון בפיתוח ביוסנסור אלקטרוני יעיל היא התכונות הפיזיקוכימיות כגון האופי המוליך למחצה והיציבות הכימית של חומר הציפוי המשמש לייצור משטח החיישן33. חומרים קונבנציונליים כמו סיליקון המשמש ל-FET הגבילו את הרגישות של חיישנים משום שהם דורשים שכבות תחמוצת דחוקות בין תעלת הטרנזיסטור לבין סביבה מסוימת לתפקוד תקיןשל 34. יתר על כן, טרנזיסטורים של סיליקון רגישים לסביבות מלח גבוהות, ובכך מקשים על מדידת אינטראקציות ביולוגיות בסביבתם הטבעית. הביוסנסור מבוסס הגרפן מוצג כחלופה מכיוון שהוא מציע יציבות כימית מעולה ושדה חשמלי. מאחר שגרפן הוא שכבה אטומית יחידה של פחמן, הוא גם רגיש ביותר כמוליכה למחצה וגם תואם כימית לתמיסות ביולוגיות; שתי התכונות הללו רצויות כדי ליצור ביוסנסורים אלקטרוניים תואמים35. פוטנציאל ההעמסה האולטרה-גבוה המדהים של ביו-מולקולות המוצע על ידי ביוסנסורים מצופים גרפן מוביל לפיתוח טכנולוגיית FEB מבוססת-גרפן.
עקרון טכנולוגיית FEB: FEB היא טכניקת זיהוי ביומולקולרית נטולת תוויות המודדת את הזרם החשמלי דרך הביו-סנסור של הגרפן שאליו משותקות מטרות הקישור. אינטראקציות בין החלבון המשותק לבין האנליט גורמות לשינויים בזרם המנוטרים בזמן אמת, ומאפשרים מדידות קינטיות מדויקות36.
מכשור: מערכת FEB כוללת שבב חיישן טרנזיסטור בעל אפקט שדה גרפן (gFET) וקורא אלקטרוני שמפעיל מתח קבוע לאורך כל הניסוי (איור 1). האנליט מיושם בתמיסה על חלבון המטרה המשותק על פני השטח של הביו-סנסור. כאשר מתרחשת אינטראקציה, שינוי בזרם נמדד ומתועד בזמן אמת. ככל שריכוז האנליטים עולה, גם החלק של האנליט המאוגד יגדל, מה שיגרום לחילופים גבוהים יותר בזרם. באמצעות תוכנת הניתוח האוטומטית המסופקת עם המכשיר (טבלת החומרים), I-Response נמדדת ומתועדת במונחים של יחידות ביוסנסינג (BU)37. I-Response מוגדר כשינוי בזרם (I) באמצעות שבב הביו-סנסור הנמדד בזמן אמת על סמך האינטראקציה של המטרה המשותקת עם האנליט. תוכנת הניתוח האוטומטית FEB יכולה לנתח הן את ה-I-Response והן את ה-C-Response לאירועי אינטראקציה דינמיים, כאשר ה-C-Response מתעד את השינויים בקיבוליות (C). הווריאציות הן ב-I-Response והן ב-C-Response מתאימות ישירות לשבריר של האנליט המאוגד וניתן לנתח אותן עוד יותר כדי ליצור ערכי KD . העדפת ברירת המחדל של תוכנת הניתוח האוטומטית היא I-Response.
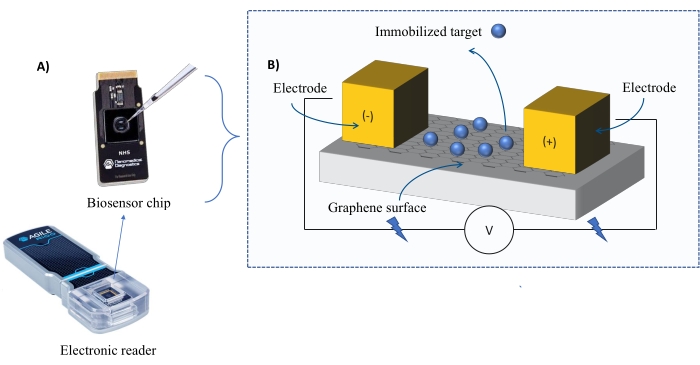
איור 1: סקירה כללית של מערך הניסוי. (B) סקירה כללית של רכיבי השבב. השבב מחובר לשתי אלקטרודות המספקות זרם למערכת. פני השטח של השבב מכוסים בגרפן, שכאשר הוא מופעל יכול לקשור את המטרה. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
מתודולוגיה:
בתחילה, שבב הביוסנסור המופעל מוכנס למכשיר FEB (איור 1) ולאחר מכן ביצוע השלבים המתוארים להלן: (1) כיול: (1) כיול: הניסוי מתחיל בכיול המערכת באמצעות 1x מי מלח עם מאגר פוספט (PBS; pH = 7.4) כדי ליצור את תגובת שיווי המשקל הבסיסית. (2) שיוך: האנליט מוצג בשבב, וה-I-Response מנוטר עד להגשמת רוויית הקשירה. (3) דיסוציאציה: האנליט מנותק באמצעות 1x PBS. (4) התחדשות: שרידי האנליט מוסרים באמצעות 1x PBS. (5) כביסה: בסך הכל חמש שטיפות מבוצעות באמצעות 1x PBS להסרה יסודית של האנליטים הכבולים והלא מאוגדים מהשבב.
ניתוח:
ניתוח הנתונים מתבצע באמצעות התוכנה האוטומטית לחלוטין המסופקת עם המכשיר. תוכנת הניתוח האוטומטית יוצרת עלילת היל פיט עם ערך KD . עלילת ההתאמה של היל מתארת את הקשר של אנליט לחלבון המטרה כפונקציה של ריכוזי אנליטים. הריכוז שבו מושגת תגובה חצי מקסימלית הוא פרופורציונלי לערך KD . ערך KD נמוך מייצג זיקת קשירה גבוהה ולהיפך.
כדי לאמת את הנתונים המתקבלים מניסוי FEB, תגובות I מופקות מכל נקודת קריאה עבור כל ריכוז אנאליטים באמצעות תוכנת סקירת/ייצוא הנתונים וניתן לייצא אותן לתוכנות ניתוח סטטיסטיות אחרות (ראה טבלת חומרים) כפי שיוסבר להלן.
