Biomolekylære interaktioner:
Proteiner er væsentlige dele af organismer og deltager i adskillige molekylære veje såsom cellemetabolisme, cellestruktur, cellesignalering, immunresponser, celleadhæsion og meget mere. Mens nogle proteiner udfører deres funktion(er) uafhængigt, interagerer de fleste proteiner med andre proteiner ved hjælp af en bindende grænseflade til at koordinere korrekt biologisk aktivitet1.
Biomolekylære interaktioner kan hovedsageligt klassificeres ud fra de forskellige strukturelle og funktionelle egenskaber ved involverede proteiner2, for eksempel baseret på proteinoverfladerne, den komplekse stabilitet eller persistensen af interaktioner3. Identifikation af essentielle proteiner og deres roller i biomolekylære interaktioner er afgørende for at forstå biokemiske mekanismer på molekylært niveau4. I øjeblikket er der forskellige tilgange til at detektere disse interaktioner5: in vitro6, in silico7, i levende celler8, ex vivo9 og in vivo10, hvor hver har sine egne styrker og svagheder.
In vivo-assays udføres ved hjælp af hele dyret som et eksperimentelt værktøj11, ogt ex vivo-assays udføres på vævsekstrakter eller hele organer (f.eks. Hjerte, hjerne, lever) i et kontrolleret eksternt miljø ved at give minimale ændringer i naturlige forhold. Den mest almindelige anvendelse af in vivo– og ex vivo-undersøgelser er at evaluere farmakokinetikken, farmakodynamikken og toksicitetsvirkningerne af potentielle farmakologiske agenser før humane forsøg ved at sikre deres samlede sikkerhed og virkning12.
Biomolekylære interaktioner kan også påvises i levende celler. Billeddannelse af levende celler giver os mulighed for at observere dynamiske interaktioner, når de udfører reaktionerne fra en bestemt biokemisk vej13. Desuden kan detektionsteknikker, såsom bioluminescens eller fluorescensresonansenergioverførsel, give information om, hvor og hvornår disse interaktioner forekommer i cellen14. Selvom detektion i levende celler tilbyder vigtige detaljer, er disse detektionsmetoder afhængige af optik og etiketter, som muligvis ikke afspejler den oprindelige biologi; de er også mindre kontrollerede end in vitro-metoder og kræver specialiseret ekspertise for at udføre15.
In silico-beregningsmetoderne anvendes primært til storstilet screening af målmolekyler før in vitro-forsøgene . Beregningsmæssige forudsigelsesmetoder, computerbaserede databaser, molekylær docking, kvantitative struktur-aktivitetsforhold og andre molekylære dynamiksimuleringsmetoder er blandt de veletablerede i silico-værktøjer 16. Sammenlignet med besværlige eksperimentelle teknikker kan in silico-værktøjerne let lave forudsigelser med høj følsomhed, men med reduceret nøjagtighed i prædiktiv ydeevne17.
In vitro-assays udføres med mikroorganismer eller biologiske molekyler uden for deres standard biologiske kontekst. At skildre biomolekylære interaktioner gennem in vitro-metoder er afgørende for at forstå proteinfunktioner og biologien bag det komplekse netværk af cellefunktion. Den foretrukne analysemetode vælges i henhold til proteinets iboende egenskaber, kinetiske værdier og interaktionsmåden og intensiteten18,19.
Hsp90/Cdc37-interaktionen:
Chaperon-kinase-vejen, der forbinder Hsp90 og Cdc37, er et lovende terapeutisk mål i tumorbiologi20. Hsp90 spiller en central rolle i cellecykluskontrol, proteinsamling, celleoverlevelse og signalveje. Proteiner, der er afhængige af Hsp90 for deres funktioner, leveres til Hsp90 til kompleksnering gennem en co-chaperon, såsom Cdc37. Hsp90/Cdc37-komplekset styrer foldningen af de fleste proteinkinaser og fungerer som et knudepunkt for en lang række intracellulære signalnetværk21. Det er et lovende antitumormål på grund af dets forhøjede ekspression i forskellige maligniteter, herunder akut myeloblastisk leukæmi, multipelt myelom og hepatocellulært carcinom22,23.
Almindeligt anvendt in vitro biomolekylære interaktionsdetekteringsteknikker
Co-immunoprecipitation (co-IP) er en teknik, der er afhængig af antigen-antistofs specificitet for at identificere biologisk relevante interaktioner24. Den primære ulempe ved denne metode er dens manglende evne til at detektere interaktioner med lav affinitet og kinetiske værdier24. Biofysiske metoder såsom isotermisk titreringskalorimetri (ITC), overfladeplasmonresonans (SPR), biolagsinterferometri (BLI) og FEB-teknologi foretrækkes til at bestemme de kinetiske værdier.
ITC er en biofysisk detektionsmetode baseret på bestemmelse af bindingsenergi sammen med en komplet termodynamikanalyse til karakterisering af biomolekylære interaktioner25. Den primære fordel ved ITC er, at det ikke kræver nogen mærkning eller fiksering af målproteinet. De største vanskeligheder, som ITC støder på, er den høje koncentration af målprotein, der kræves til et eksperiment, og vanskeligheden ved at analysere ikke-kovalente komplekser på grund af små bindende entalpier26. Både SPR og BLI er etiketfrie biofysiske teknikker, der er afhængige af immobilisering af målmolekylet på sensoroverfladen efterfulgt af efterfølgende injektioner af analytten over det immobiliserede mål27,28. I SPR måles ændringer i brydningsindekset under biomolekylære interaktioner27; i BLI registreres interferensen i reflekteret lys i realtid som en ændring i bølgelængden som en funktion af tid28. Både SPR og BLI deler fælles fordele ved at tilbyde høj specificitet, følsomhed og detektionsfunktioner29. I begge metoder immobiliseres målproteinet på biosensoroverflader, og der kan derfor være et vist tab af målets oprindelige konformation, hvilket gør det vanskeligt at skelne mellem specifikke vs. ikke-specifikke interaktioner30. BLI bruger dyre engangsfiberoptiske biosensorer til at immobilisere målet, og er derfor en dyr teknik31. Sammenlignet med disse veletablerede biomolekylære detektionsværktøjer tilbyder FEB-teknologien en pålidelig og etiketfri platform ved at bruge lave nanomolære koncentrationer til biomolekylær detektion i realtid med kinetisk karakterisering. FEB-teknologien overvinder også de boblende udfordringer, som ITC står over for, og er mere omkostningseffektiv sammenlignet med SPR eller BLI.
De felteffekttransistorbaserede biosensorer (FET) er et voksende felt til påvisning af biomolekylære interaktioner ved at tilbyde forskellige biomedicinske applikationer. I FET-systemet immobiliseres målene til biosensorchipsene, og interaktioner detekteres ved ændringer i konduktans32. Det unikke træk, der skal tages i betragtning ved udviklingen af en effektiv elektronisk biosensor, er de fysisk-kemiske egenskaber såsom den halvledende karakter og kemiske stabilitet af det belægningsmateriale, der anvendes til fremstilling af sensoroverfladen33. Konventionelle materialer som silicium, der anvendes til FET, har begrænset sensorernes følsomhed, fordi det kræver oxidlag klemt inde mellem transistorkanalen og et specifikt miljø for korrekt funktion34. Desuden er siliciumtransistorer følsomme over for miljøer med højt saltindhold, hvilket gør det svært at måle biologiske interaktioner i deres naturlige miljø. Den grafenbaserede biosensor præsenteres som et alternativ, da den tilbyder fremragende kemisk stabilitet og elektrisk felt. Da grafen er et enkelt atomlag af kulstof, er det både ekstremt følsomt som en halvleder og kemisk kompatibelt med biologiske opløsninger; begge disse kvaliteter er ønskelige for at generere kompatible elektroniske biosensorer35. Det bemærkelsesværdige ultrahøje belastningspotentiale i biomolekyler, der tilbydes af grafenbelagte biosensorer, fører til udviklingen af grafenbaserede biosensorer FEB-teknologi.
Princippet om FEB-teknologi: FEB er en etiketfri biomolekylær detektionsteknik, der måler den elektriske strøm gennem grafenbiosensoren, som bindingsmålene immobiliseres til. Interaktioner mellem det immobiliserede protein og analytten resulterer i ændringer i strømmen, der overvåges i realtid, hvilket muliggør nøjagtige kinetiske målinger36.
Instrumentering: FEB-systemet består af en grafenfelteffekttransistor (gFET) sensorchip og en elektronisk læser, der anvender en konstant spænding gennem hele eksperimentet (figur 1). Analytten påføres i opløsning på målproteinet immobiliseret på biosensoroverfladen. Når en interaktion opstår, måles og registreres en ændring i strømmen i realtid. Efterhånden som analytkoncentrationen stiger, vil fraktionen af bundet analyt også stige, hvilket forårsager højere vekslinger i strømmen. Ved hjælp af den automatiserede analysesoftware, der følger med instrumentet (Table of Materials), måles og registreres I-Response i form af biosensingenheder (BU)37. I-Response defineres som ændringen i strømmen (I) gennem biosensorchippen målt i realtid ved interaktionen mellem det immobiliserede mål og analytten. DEN FEB automatiserede analysesoftware kan analysere både I-Response og C-Response til dynamiske interaktionshændelser, hvor C-Response registrerer ændringerne i kapacitansen (C). Variationerne i både I-Response og C-Response svarer direkte til brøkdelen af bundet analyt og kan analyseres yderligere for at generere KD-værdier . Den automatiserede analysesoftwares standardpræference er I-Response.
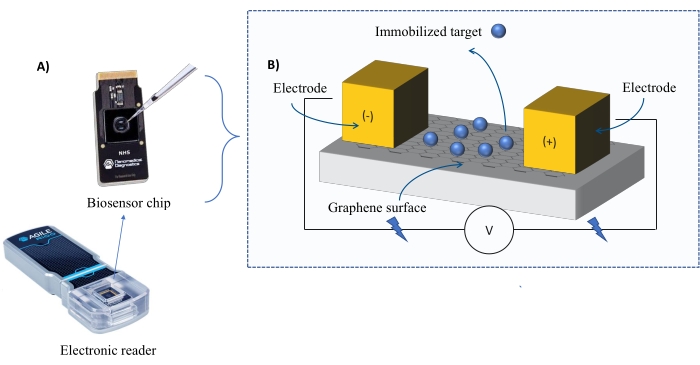
Figur 1: Oversigt over forsøgsopstillingen. (A) Grafenbaseret chip og elektronisk læser. (B) En oversigt over chipkomponenterne. Chippen er fastgjort til to elektroder, der leverer strøm til systemet. Overfladen af chippen er dækket af grafen, som, når den aktiveres, kan binde målet. Klik her for at se en større version af denne figur.
Metodik:
Indledningsvis indsættes den aktiverede biosensorchip i FEB-enheden (figur 1) efterfulgt af udførelsen af nedenstående trin: (1) Kalibrering: Eksperimentet starter med systemkalibrering ved anvendelse af 1x fosfatbufret saltvand (PBS; pH = 7,4) for at skabe baseline-ligevægtsresponsen. (2) Tilknytning: Analytten indføres i chippen, og I-Response overvåges, indtil bindingsmætningen er nået. (3) Dissociation: Analytten dissocieres ved hjælp af 1x PBS. (4) Regenerering: Rester af analytten fjernes ved hjælp af 1x PBS. (5) Vask: I alt fem vaske udføres ved hjælp af 1x PBS til grundig fjernelse af de bundne og ubundne analytter fra chippen.
Analyse:
Dataanalyse udføres ved hjælp af den fuldautomatiske software, der følger med instrumentet. Den automatiserede analysesoftware genererer et Hill fit-plot med en KD-værdi . Hill fit-plottet beskriver en analyts tilknytning til målproteinet som en funktion af analytkoncentrationer. Koncentrationen, ved hvilken der opnås et halvt maksimalt respons, er proportional med KD-værdien . En lav KD-værdi repræsenterer høj bindingsaffinitet og omvendt.
For at validere de data, der er opnået fra FEB-eksperimentet, ekstraheres I-Responses fra hvert udlæsningspunkt for hver analytkoncentration ved hjælp af datagennemgangs-/eksportsoftwaren og kan eksporteres til anden statistisk analysesoftware (se materialetabellen) som forklaret nedenfor.
