Interações biomoleculares:
As proteínas são partes essenciais dos organismos e participam de inúmeras vias moleculares, como metabolismo celular, estrutura celular, sinalização celular, respostas imunes, adesão celular e muito mais. Enquanto algumas proteínas executam suas funções de forma independente, a maioria das proteínas interagem com outras proteínas usando uma interface de ligação para coordenar a atividade biológica adequada1.
As interações biomoleculares podem ser classificadas principalmente com base nas distintas características estruturais e funcionais das proteínas envolvidas2, por exemplo, com base nas superfícies proteicas, na estabilidade complexa ou na persistência das interações3. Identificar proteínas essenciais e seus papéis nas interações biomoleculares é vital para a compreensão de mecanismos bioquímicos no nível molecular4. Atualmente, existem várias abordagens para detectar essas interações5: in vitro6, no silico7, nas células vivas8, ex vivo9, e in vivo10 com cada um tendo seus próprios pontos fortes e fracos.
Os ensaios in vivo são realizados utilizando todo o animal como ferramenta experimental11, e os ensaiost he ex vivo são realizados em extratos de tecido ou órgãos inteiros (por exemplo, coração, cérebro, fígado) em um ambiente externo controlado, proporcionando alterações mínimas em condições naturais. A aplicação mais comum de estudos in vivo e ex vivo é avaliar a farmacocinética, a farmacodinâmica e os efeitos de toxicidade de potenciais agentes farmacológicos antes dos testes em humanos, garantindo sua segurança e eficácia globais12.
Interações biomoleculares também podem ser detectadas dentro de células vivas. As células vivas de imagem nos permitem observar interações dinâmicas enquanto executam as reações de uma determinada via bioquímica13. Além disso, técnicas de detecção, como bioluminescência ou transferência de ressonância fluorescência, podem fornecer informações sobre onde e quando essas interações ocorrem dentro da célula14. Embora a detecção em células vivas ofereça detalhes cruciais, essas metodologias de detecção dependem de óptica e rótulos, que podem não refletir a biologia nativa; eles também são menos controlados do que os métodos in vitro e requerem especialização especializada para realizar15.
Os métodos computacionais em silico são usados principalmente para a triagem em larga escala de moléculas-alvo antes dos experimentos in vitro . Métodos de previsão computacional, bancos de dados baseados em computador, acoplamento molecular, relações de estrutura-atividade quantitativa e outras abordagens de simulação de dinâmica molecular estão entre os bem estabelecidos nas ferramentas silico 16. Em comparação com técnicas experimentais laboriosas, as ferramentas em silico podem facilmente fazer previsões com alta sensibilidade, mas com menor precisão no desempenho preditivo17.
Ensaios in vitro são realizados com microrganismos ou moléculas biológicas fora de seu contexto biológico padrão. Retratar interações biomoleculares através de métodos in vitro é fundamental para entender as funções proteicas e a biologia por trás da complexa rede de funcionamento celular. A metodologia de ensaio preferencial é escolhida de acordo com as propriedades intrínsecas da proteína, valores cinéticos e o modo e intensidade das interações18,19.
A interação Hsp90/Cdc37:
A via acompanhante-quinase, que conecta Hsp90 e Cdc37, é um alvo terapêutico promissor na biologia tumoral20. O Hsp90 desempenha um papel central no controle do ciclo celular, montagem de proteínas, sobrevivência celular e caminhos de sinalização. Proteínas que dependem do Hsp90 para suas funções são entregues ao Hsp90 para complexação através de um co-acompanhante, como o Cdc37. O complexo Hsp90/Cdc37 controla a dobra da maioria das quinases proteicas e serve como um hub para uma infinidade de redes de sinalização intracelular21. É um alvo anti-tumor promissor devido à sua expressão elevada em várias malignidades, incluindo leucemia mieloblástica aguda, mieloma múltiplo e carcinoma hepatocelular22,23.
Comumente usadas técnicas de detecção de interação biomolecular in vitro
A co-imunoprecipitação (co-IP) é uma técnica que depende da especificidade do anticorpo de antígeno para identificar interações biologicamente relevantes24. A principal desvantagem deste método é sua incapacidade de detectar interações de baixa afinidade e valores cinéticos24. Métodos biofísicos como calorimetria de titulação isotérmica (ITC), ressonância de plasmon superficial (SPR), interferometria biocamida (BLI) e tecnologia FEB são preferidos para determinar os valores cinéticos.
ITC é um método de detecção biofísica baseado na determinação da energia vinculante, juntamente com uma análise termodinâmica completa para caracterizar interações biomoleculares25. A principal vantagem do ITC é que ele não requer nenhuma rotulagem ou fixação da proteína alvo. As principais dificuldades encontradas pelo ITC são a alta concentração de proteína-alvo necessária para um experimento e a dificuldade em analisar complexos não covalentes devido a pequenas indeseçõesde ligação 26. Tanto o SPR quanto o BLI são técnicas biofísicas sem rótulos que dependem da imobilização da molécula alvo na superfície do sensor, seguidas de injeções subsequentes do analito sobre o alvo imobilizado27,28. Em SPR, são medidas alterações no índice refrativo durante interações biomoleculares27; em BLI, a interferência na luz refletida é registrada em tempo real como uma mudança no comprimento de onda em função do tempo28. Tanto o SPR quanto o BLI compartilham vantagens comuns de oferecer altas especificidades, sensibilidade e recursos de detecção29. Em ambos os métodos, a proteína alvo é imobilizada em superfícies biosensoras e, portanto, pode haver alguma perda da conformação nativa do alvo, o que dificulta a discriminação entre interações específicas versus não específicas30. A BLI usa biosensores descartáveis de fibra óptica caros para imobilizar o alvo e, portanto, é uma técnicacara 31. Em comparação com essas ferramentas bem estabelecidas de detecção biomolecular, a tecnologia FEB oferece uma plataforma confiável e livre de rótulos usando baixas concentrações de nanomolar para detecção biomolecular em tempo real com caracterização cinética. A tecnologia FEB também supera os desafios borbulhantes enfrentados no ITC e é mais econômica em comparação com a SPR ou BLI.
O biosensores baseados em transistor de efeito de campo (FET) é um campo emergente para detectar interações biomoleculares, oferecendo aplicações biomédicas variadas. No sistema FET, os alvos são imobilizados para os chips biosensor e as interações são detectadas por alterações na condutância32. A característica única a ser considerada no desenvolvimento de um biosensor eletrônico eficiente são as propriedades físico-químicas, como a natureza semi-condutora e a estabilidade química do material de revestimento usado para fabricar a superfície do sensor33. Materiais convencionais como o silício usado para fet limitaram a sensibilidade dos sensores porque requer camadas de óxido entre o canal do transistor e um ambiente específico para o bom funcionamento34. Além disso, os transistores de silício são sensíveis a ambientes de alto sal, dificultando a mensuração das interações biológicas em seu ambiente natural. O biosensor à base de grafeno é apresentado como uma alternativa, pois oferece excelente estabilidade química e campo elétrico. Uma vez que o grafeno é uma única camada atômica de carbono, é extremamente sensível como um semicondutor e quimicamente compatível com soluções biológicas; ambas as qualidades são desejáveis para gerar biosensores eletrônicos compatíveis35. O notável potencial de carregamento ultra-alto de biomoléculas oferecido por biosensores revestidos de grafeno leva ao desenvolvimento da tecnologia FEB baseada em grafeno.
Princípio da tecnologia FEB: FEB é uma técnica de detecção biomolecular sem rótulos que mede a corrente elétrica através do biosensor de grafeno ao qual os alvos de ligação são imobilizados. As interações entre a proteína imobilizada e o analito resultam em alterações na corrente que são monitoradas em tempo real, permitindo medições cinéticas precisas36.
Instrumentação: O sistema FEB compreende um chip de sensor transistor de efeito de campo de grafeno (gFET) e um leitor eletrônico que aplica uma tensão constante ao longo do experimento (Figura 1). O analito é aplicado em solução para a proteína alvo imobilizada na superfície do biosensor. Quando ocorre uma interação, uma alteração na corrente é medida e registrada em tempo real. À medida que a concentração de analito aumenta, a fração de analito ligado também aumentará, causando maiores alterações na correnteza. Utilizando o software de análise automatizada fornecido com o instrumento (Tabela de Materiais), o I-Response é medido e registrado em termos de unidades de biosensação (BU)37. A I-Resposta é definida como a alteração na corrente (I) através do chip biosensor medido em tempo real após a interação do alvo imobilizado com o analito. O software de análise automatizada FEB pode analisar tanto o I-Response quanto o C-Response para eventos de interação dinâmica, onde o C-Response registra as alterações na capacitância (C). As variações tanto na I-Resposta quanto na Resposta C correspondem diretamente à fração de analito vinculado e podem ser analisadas para gerar valores KD . A preferência padrão do software de análise automatizada é I-Response.
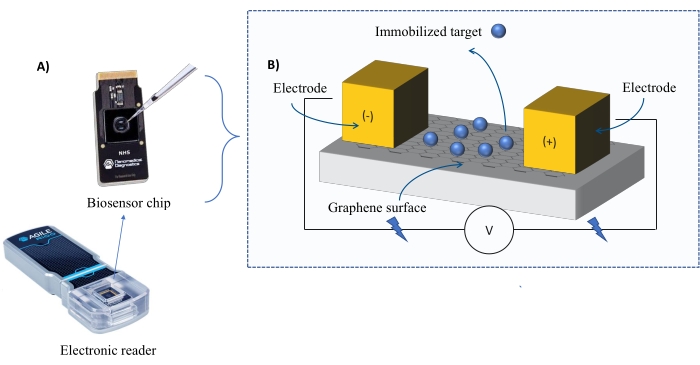
Figura 1: Visão geral da configuração experimental. (A) chip baseado em grafeno e um leitor eletrônico. (B) Uma visão geral dos componentes do chip. O chip está ligado a dois eletrodos que fornecem corrente ao sistema. A superfície do chip é coberta com grafeno, que quando ativado pode ligar o alvo. Clique aqui para ver uma versão maior desta figura.
Metodologia:
Inicialmente, o chip biosensor ativado é inserido no dispositivo FEB (Figura 1) seguido da execução das etapas abaixo: (1) Calibração: O experimento começa com a calibração do sistema usando 1x salina tamponada de fosfato (PBS; pH = 7,4) para criar a resposta de equilíbrio de linha de base. (2) Associação: O analito é introduzido no chip e o I-Response é monitorado até que a saturação de vinculação seja alcançada. (3) Dissociação: O analito é dissociado utilizando 1x PBS. (4) Regeneração: Os remanescentes do analito são removidos utilizando-se 1x PBS. (5) Lavagem: Um total de cinco lavagens são realizadas utilizando 1x PBS para a remoção completa dos analitos amarrados e desvinculados do chip.
Análise:
A análise de dados é realizada utilizando-se o software totalmente automatizado fornecido com o instrumento. O software de análise automatizada gera um gráfico de ajuste hill com um valor KD . O enredo de ajuste de Hill descreve a associação de um analito à proteína alvo em função de concentrações de analitos. A concentração na qual uma resposta semi-máxima é alcançada é proporcional ao valor KD . Um baixo valor KD representa alta afinidade vinculante e vice-versa.
Para validar os dados obtidos a partir do experimento FEB, as Respostas I são extraídas de cada ponto de leitura para cada concentração de analitos usando o software de revisão/exportação de dados e podem ser exportadas para outros softwares de análise estatística (ver Tabela de Materiais), conforme explicado abaixo.
