Interacciones biomoleculares:
Las proteínas son partes esenciales de los organismos y participan en numerosas vías moleculares como el metabolismo celular, la estructura celular, la señalización celular, las respuestas inmunes, la adhesión celular y más. Mientras que algunas proteínas realizan su(s) función(es) de forma independiente, la mayoría de las proteínas interactúan con otras proteínas utilizando una interfaz de unión para coordinar la actividad biológica adecuada1.
Las interacciones biomoleculares se pueden clasificar principalmente en función de las distintas características estructurales y funcionales de las proteínas involucradas2, por ejemplo, en función de las superficies de las proteínas, la estabilidad compleja o la persistencia de las interacciones3. La identificación de proteínas esenciales y su papel en las interacciones biomoleculares es vital para comprender los mecanismos bioquímicos a nivel molecular4. Actualmente, existen varios enfoques para detectar estas interacciones5: in vitro6, in silico7, en células vivas8, ex vivo9 e in vivo10, cada una con sus propias fortalezas y debilidades.
Los ensayos in vivo se realizan utilizando todo el animal como herramienta experimental11, ylos ensayos ex vivo se realizan en extractos de tejidos u órganos enteros (por ejemplo, corazón, cerebro, hígado) en un entorno externo controlado proporcionando alteraciones mínimas en condiciones naturales. La aplicación más común de los estudios in vivo y ex vivo es evaluar los efectos farmacocinéticos, farmacodinámicos y de toxicidad de los posibles agentes farmacológicos antes de los ensayos en humanos garantizando su seguridad y eficaciagenerales 12.
Las interacciones biomoleculares también se pueden detectar dentro de las células vivas. Las imágenes de células vivas nos permiten observar interacciones dinámicas a medida que ejecutan las reacciones de una vía bioquímica particular13. Además, las técnicas de detección, como la bioluminiscencia o la transferencia de energía de resonancia de fluorescencia, pueden proporcionar información sobre dónde y cuándo ocurren estas interacciones dentro de la célula14. Aunque la detección en células vivas ofrece detalles cruciales, estas metodologías de detección se basan en ópticas y etiquetas, que pueden no reflejar la biología nativa; también están menos controlados que los métodos in vitro y requieren experiencia especializada para realizar15.
Los métodos computacionales in silico se utilizan principalmente para el cribado a gran escala de moléculas diana antes de los experimentos in vitro . Los métodos de predicción computacional, las bases de datos basadas en computadora, el acoplamiento molecular, las relaciones cuantitativas estructura-actividad y otros enfoques de simulación de dinámica molecular se encuentran entre las herramientas in silico bien establecidas16. En comparación con las laboriosas técnicas experimentales, las herramientas in silico pueden hacer predicciones fácilmente con alta sensibilidad, pero con una precisión reducida en el rendimiento predictivo17.
Los ensayos in vitro se realizan con microorganismos o moléculas biológicas fuera de su contexto biológico estándar. Retratar las interacciones biomoleculares a través de métodos in vitro es fundamental para comprender las funciones de las proteínas y la biología detrás de la compleja red de funcionamiento celular. La metodología de ensayo preferida se elige de acuerdo con las propiedades intrínsecas de la proteína, los valores cinéticos y el modo e intensidad de las interacciones18,19.
La interacción Hsp90/Cdc37:
La vía chaperona-quinasa, que conecta Hsp90 y Cdc37, es una diana terapéutica prometedora en biología tumoral20. Hsp90 desempeña un papel central en el control del ciclo celular, el ensamblaje de proteínas, la supervivencia celular y las vías de señalización. Las proteínas que dependen de Hsp90 para sus funciones se entregan a Hsp90 para su complejación a través de una co-chaperona, como Cdc37. El complejo Hsp90/Cdc37 controla el plegamiento de la mayoría de las proteínas quinasas y sirve como centro para una multitud de redes de señalización intracelular21. Es una diana antitumoral prometedora debido a su elevada expresión en diversas neoplasias malignas, incluyendo leucemia mieloblástica aguda, mieloma múltiple y carcinoma hepatocelular22,23.
Técnicas de detección de interacción biomolecular in vitro de uso común
La coinmunoprecipitación (co-IP) es una técnica que se basa en la especificidad antígeno-anticuerpo para identificar interacciones biológicamente relevantes24. La principal desventaja de este método es su incapacidad para detectar interacciones de baja afinidad y valores cinéticos24. Se prefieren métodos biofísicos como la calorimetría de titulación isotérmica (ITC), la resonancia de plasmón de superficie (SPR), la interferometría de biocapa (BLI) y la tecnología FEB para determinar los valores cinéticos.
ItC es un método de detección biofísica basado en la determinación de la energía de unión junto con un análisis termodinámico completo para caracterizar las interacciones biomoleculares25. La principal ventaja de ITC es que no requiere ningún etiquetado o fijación de la proteína objetivo. Las principales dificultades encontradas por el ITC son la alta concentración de proteína objetivo requerida para un experimento y la dificultad para analizar complejos no covalentes debido a pequeñas entalpías de unión26. Tanto SPR como BLI son técnicas biofísicas sin etiquetas que se basan en la inmovilización de la molécula diana en la superficie del sensor, seguida de inyecciones posteriores del analito sobre el objetivo inmovilizado27,28. En SPR se miden alteraciones en el índice de refracción durante las interacciones biomoleculares27; en BLI, la interferencia en la luz reflejada se registra en tiempo real como un cambio en la longitud de onda en función del tiempo28. Tanto SPR como BLI comparten ventajas comunes de ofrecer capacidades de alta especificidad, sensibilidad y detección29. En ambos métodos, la proteína diana se inmoviliza en las superficies de los biosensores y, por lo tanto, puede haber cierta pérdida de la conformación nativa del objetivo, lo que dificulta la discriminación entre interacciones específicas vs. no específicas30. BLI utiliza costosos biosensores desechables de fibra óptica para inmovilizar el objetivo y, por lo tanto, es una técnica costosa31. En comparación con estas herramientas de detección biomolecular bien establecidas, la tecnología FEB ofrece una plataforma confiable y libre de etiquetas mediante el uso de bajas concentraciones nanomolares para la detección biomolecular en tiempo real con caracterización cinética. La tecnología FEB también supera los burbujeantes desafíos que enfrenta el ITC y es más rentable en comparación con SPR o BLI.
Los biosensores basados en transistores de efecto de campo (FET) son un campo emergente para detectar interacciones biomoleculares al ofrecer diversas aplicaciones biomédicas. En el sistema FET, los objetivos se inmovilizan a los chips del biosensor y las interacciones se detectan mediante cambios en la conductancia32. La característica única a considerar en el desarrollo de un biosensor electrónico eficiente son las propiedades fisicoquímicas como la naturaleza semiconductora y la estabilidad química del material de recubrimiento utilizado para fabricar la superficie del sensor33. Los materiales convencionales como el silicio utilizado para FET han limitado la sensibilidad de los sensores porque requiere capas de óxido intercaladas entre el canal del transistor y un entorno específico para un funcionamiento adecuado34. Además, los transistores de silicio son sensibles a ambientes con alto contenido de sal, lo que dificulta la medición de las interacciones biológicas en su entorno natural. El biosensor basado en grafeno se presenta como una alternativa, ya que ofrece una excelente estabilidad química y campo eléctrico. Dado que el grafeno es una sola capa atómica de carbono, es extremadamente sensible como semiconductor y químicamente compatible con soluciones biológicas; ambas cualidades son deseables para generar biosensores electrónicos compatibles35. El notable potencial de carga ultra alta de las biomoléculas ofrecidas por los biosensores recubiertos de grafeno conduce al desarrollo de la tecnología FEB de biosensores basados en grafeno.
Principio de la tecnología FEB: FEB es una técnica de detección biomolecular sin etiqueta que mide la corriente eléctrica a través del biosensor de grafeno al que se inmovilizan los objetivos de unión. Las interacciones entre la proteína inmovilizada y el analito dan lugar a alteraciones en la corriente que se monitorizan en tiempo real, lo que permite mediciones cinéticas precisas36.
Instrumentación: El sistema FEB comprende un chip sensor de transistor de efecto de campo de grafeno (gFET) y un lector electrónico que aplica un voltaje constante durante todo el experimento (Figura 1). El analito se aplica en solución a la proteína diana inmovilizada en la superficie del biosensor. Cuando se produce una interacción, se mide y registra una alteración en la corriente en tiempo real. A medida que aumenta la concentración de analitos, la fracción de analito unido también aumentará, causando mayores alteraciones en la corriente. Utilizando el software de análisis automatizado proporcionado con el instrumento (Tabla de Materiales), I-Response se mide y registra en términos de unidades de biodetección (BU)37. I-Response se define como la alteración en la corriente (I) a través del chip biosensor medida en tiempo real sobre la interacción del objetivo inmovilizado con el analito. El software de análisis automatizado FEB puede analizar tanto la I-Response como la C-Response a eventos de interacción dinámica, donde la C-Response registra las alteraciones en la capacitancia (C). Las variaciones tanto en la I-Response como en la C-Response corresponden directamente a la fracción de analito unido y pueden analizarse más a fondo para generar valores KD . La preferencia predeterminada del software de análisis automatizado es I-Response.
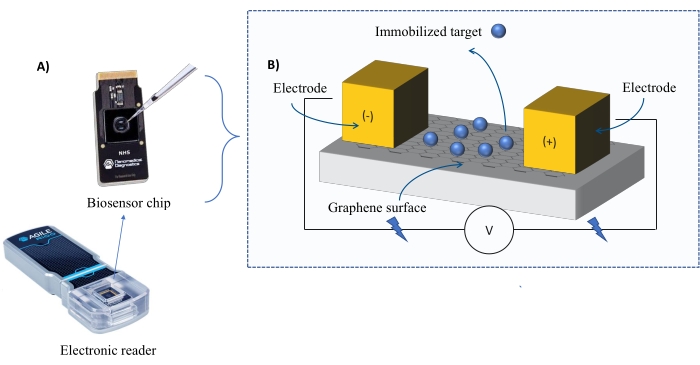
Figura 1: Descripción general de la configuración experimental. (A) Chip basado en grafeno y un lector electrónico. (B) Una visión general de los componentes del chip. El chip está unido a dos electrodos que suministran corriente al sistema. La superficie del chip está cubierta con grafeno, que cuando se activa puede unir al objetivo. Haga clic aquí para ver una versión más grande de esta figura.
Metodología:
Inicialmente, el chip biosensor activado se inserta en el dispositivo FEB (Figura 1) seguido de la ejecución de los pasos descritos a continuación: (1) Calibración: El experimento comienza con la calibración del sistema utilizando 1x solución salina tamponada con fosfato (PBS; pH = 7.4) para crear la respuesta de equilibrio basal. (2) Asociación: El analito se introduce en el chip, y el I-Response se monitorea hasta que se alcanza la saturación de unión. (3) Disociación: El analito se disocia usando 1x PBS. (4) Regeneración: Los restos del analito se eliminan usando 1x PBS. (5) Lavado: Se realizan un total de cinco lavados utilizando 1x PBS para la eliminación completa de los analitos unidos y no unidos del chip.
Análisis:
El análisis de datos se realiza utilizando el software totalmente automatizado proporcionado con el instrumento. El software de análisis automatizado genera un diagrama de ajuste de Hill con un valor KD . La gráfica de ajuste de Hill describe la asociación de un analito a la proteína objetivo en función de las concentraciones de analitos. La concentración a la que se alcanza una respuesta medio máxima es proporcional al valor KD . Un valor bajo de KD representa una alta afinidad de unión y viceversa.
Para validar los datos obtenidos del experimento FEB, las respuestas I se extraen de cada punto de lectura para cada concentración de analito utilizando el software de revisión / exportación de datos y se pueden exportar a otro software de análisis estadístico (consulte la Tabla de materiales) como se explica a continuación.
