נוירו-פרוטזות חזותיות הן קבוצה של מכשירים המספקים גירויים (חשמליים, אור וכו ‘) לתאים העצביים במסלול הראייה כדי ליצור פוספנים או תחושה של ראיית האור. זוהי אסטרטגיית טיפול שנמצאת בשימוש קליני כבר כמעט עשור עבור אנשים עם עיוורון קבוע הנגרם על ידי מחלות רשתית ניווניות. בדרך כלל, מערכת שלמה תכלול מצלמה חיצונית הלוכדת את המידע החזותי סביב המשתמש, ספק כוח ויחידת מחשוב לעיבוד ותרגום התמונה לסדרה של פולסים חשמליים, ומערך אלקטרודות מושתל המממשק את הרקמה העצבית ומעביר את הפולסים החשמליים לתאים העצביים. עקרון העבודה מאפשר למקם נוירופרוסטזה חזותית באתרים שונים לאורך מסלול הראייה מהרשתית לקליפת המוח הראייתית, כל עוד היא נמצאת במורד הזרם מהרקמה הפגועה. רוב המחקרים הנוכחיים בנוירו-פרוסטזות חזותיות מתמקדים בהגברת היעילות של הגירוי ושיפור החדות המרחבית כדי לספק ראייה טבעית יותר.
במאמצים לשפר את יעילות הגירוי, מידול חישובי היה שיטה חסכונית וחסכונית בזמן כדי לאמת תכנון תותבת ולדמות את התוצאה החזותית שלה. מודלים חישוביים בתחום זה צברו פופולריות מאז 1999 כאשר גרינברג1 מודל את התגובה של תא גנגליון רשתית לגירויים חשמליים חוץ-תאיים. מאז, מודלים חישוביים שימשו כדי לייעל את הפרמטרים של הפולס החשמלי 2,3 או את העיצוב הגיאומטרי של האלקטרודה 4,5. למרות השונות במורכבות ובשאלות המחקר, מודלים אלה פועלים על ידי קביעת חלוקת המתח החשמלי בתווך (למשל, רקמה עצבית) והערכת התגובה החשמלית שהנוירונים בסביבה ייצרו עקב המתח החשמלי.
ניתן למצוא את חלוקת המתח החשמלי במוליך על ידי פתרון משוואות פואסון6 בכל המקומות:
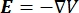
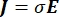
כאשר E הוא השדה החשמלי, V הוא הפוטנציאל החשמלי, J צפיפות הזרם, ו- σ היא המוליכות החשמלית. במשוואה 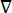 מציין אופרטור הדרגתי. במקרה של זרם נייח, תנאי הגבול הבאים מוטלים על המודל:
מציין אופרטור הדרגתי. במקרה של זרם נייח, תנאי הגבול הבאים מוטלים על המודל:
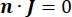
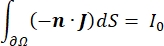
כאשר n הוא הנורמלי לפני השטח, Ω מייצג את הגבול, ו-I0 מייצג את הזרם הספציפי. יחד, הם יוצרים בידוד חשמלי בגבולות החיצוניים ויוצרים מקור זרם לגבול שנבחר. אם נניח מקור נקודה מונופולרית בתווך הומוגני בעל מוליכות איזוטרופית, ניתן לחשב את הפוטנציאל החשמלי החוץ-תאי במיקום שרירותי על ידי7:
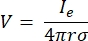
כאשר Ie הוא הזרם ו- הוא המרחק בין האלקטרודה לנקודת המדידה. כאשר התווך אינו הומוגני או אניזוטרופי, או שלמערך האלקטרודות יש אלקטרודות מרובות, חבילה חישובית לפתרון מספרי של המשוואות יכולה להיות נוחה. תוכנת מידול אלמנטים סופיים6 מפרקת את מוליך הנפח למקטעים קטנים המכונים ‘אלמנטים’. האלמנטים קשורים זה בזה כך שהשפעות השינוי באלמנט אחד משפיעות על השינוי באחרים, והוא פותר את המשוואות הפיזיקליות המשמשות לתיאור אלמנטים אלה. עם המהירות החישובית הגוברת של מחשבים מודרניים, תהליך זה יכול להסתיים תוך שניות. לאחר חישוב הפוטנציאל החשמלי, ניתן להעריך את התגובה החשמלית של תא העצב.
נוירון שולח ומקבל מידע בצורה של אותות חשמליים. אותות כאלה מגיעים בשתי צורות – פוטנציאלים מדורגים ופוטנציאלי פעולה. פוטנציאלים מדורגים הם שינויים זמניים בפוטנציאל הממברנה שבהם המתח על פני הממברנה הופך חיובי יותר (דה-פולריזציה) או שלילי יותר (היפרפולריזציה). לפוטנציאלים מדורגים יש בדרך כלל השפעות מקומיות. בתאים שמייצרים אותם, פוטנציאל פעולה הוא תגובות של הכל או לא כלום שיכולות לנוע למרחקים ארוכים לאורך האקסון. הן פוטנציאל מדורג והן פוטנציאל פעולה רגישים לסביבה החשמלית כמו גם הכימית. ספייק פוטנציאלי פעולה יכול להיווצר על ידי סוגי תאים עצביים שונים, כולל תאי גנגליון הרשתית, כאשר פוטנציאל טרנס-ממברנה סף נחצה. פוטנציאל הפעולה מזנק ומתפשט ואז מפעילים העברה סינפטית של אותות לנוירונים במורד הזרם. ניתן לעצב נוירון ככבל המחולק למקטעים גליליים, כאשר לכל מקטע יש קיבוליות והתנגדות עקב הממברנה הדו-שכבתית השומנית8. תוכנית חישובית נוירונים9 יכולה להעריך את הפעילות החשמלית של תא מעורר חשמלית על ידי הפרדת התא למספר תאים ופתרון המודל המתמטי10:
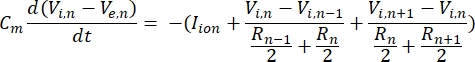
במשוואה זו, Cmהוא קיבוליות הממברנה, V e,n הוא הפוטנציאל החוץ-תאי בצומת n, Vi,n הפוטנציאל התוך-תאי בצומת n, R n ההתנגדות התוך-תאית (אורכית) בצומת n, ויון I הוא הזרם היוני העובר דרך תעלות היונים בצומת n. הערכים של V ממודל FEM מיושמים כ– Ve,n עבור כל הצמתים בנוירון כאשר הגירוי פעיל.
ניתן למדל את הזרמים הטרנס-ממברניים מתעלות יונים באמצעות נוסחאות הודג’קין-האקסלי11:
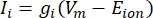
כאשר g i הוא המוליכות הספציפית של התעלה, V m פוטנציאל הטרנסממברנה (V i,n – Ve,n) ויון E פוטנציאל ההיפוך של תעלת היונים. עבור ערוצים מגודרים במתח, כגון ערוץ Na, מוצגים פרמטרים חסרי ממדים, m ו– h, המתארים את ההסתברות לפתיחה או לסגירה של הערוצים:
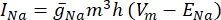
כאשר 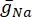 היא מוליכות הממברנה המקסימלית עבור תעלת היונים המסוימת, וערכי הפרמטרים m ו-h מוגדרים על ידי משוואות דיפרנציאליות:
היא מוליכות הממברנה המקסימלית עבור תעלת היונים המסוימת, וערכי הפרמטרים m ו-h מוגדרים על ידי משוואות דיפרנציאליות:
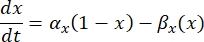
כאשר α x ו- βx הן פונקציות תלויות מתח המגדירות את קבועי הקצב של תעלת היונים. הם בדרך כלל לובשים את הטופס:
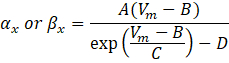
הערכים של הפרמטרים במשוואות אלה, כולל מוליכות מקסימלית, כמו גם הקבועים A, B, C ו- D, נמצאו בדרך כלל ממדידות אמפיריות.
בעזרת אבני בניין אלה, ניתן לבנות מודלים בעלי מורכבויות שונות על ידי ביצוע השלבים המתוארים. תוכנת FEM שימושית כאשר לא ניתן לפתור את משוואת פואסון באופן אנליטי, כגון במקרה של מוליכות אי-הומוגנית או אניזוטרופית במוליך הנפח או כאשר הגאומטריה של מערך האלקטרודות מורכבת. לאחר שערכי הפוטנציאל החוץ-תאי נפתרו, ניתן לפתור את מודל כבל הנוירונים באופן מספרי בתוכנה החישובית של תא העצב. שילוב שתי התוכנות מאפשר חישוב של תא עצב מורכב או רשת לשדה חשמלי לא אחיד.
מודל דו-שלבי פשוט של תא גנגליון ברשתית תחת גירוי על-טבעי ייבנה באמצעות התוכניות הנ”ל. במחקר זה, תא הגנגליון ברשתית יהיה נתון לטווח של סדרי גודל של פולסים של זרם חשמלי. גם מיקום התא ביחס לגירוי משתנה כדי להראות את הקשר בין סף המרחק. יתר על כן, המחקר כולל אימות של התוצאה החישובית כנגד מחקר in vivo של סף ההפעלה בקליפת המוח תוך שימוש בגדלים שונים של אלקטרודת גירוי12, וכן מחקר במבחנה המראה את הקשר בין מרחק האלקטרודה-נוירון לבין סף ההפעלה13.
