A growing body of work suggests that biomolecular condensates play an important role in cellular organization and numerous cellular functions, e.g., transcriptional regulation1,2,3,4,5, DNA damage repair6,7,8, chromatin organization9,10,11,12, X-chromosome inactivation13,14,15, and intracellular signaling16,17,18. In addition, the dysregulation of biomolecular condensates is implicated in many diseases, including cancers19,20,21 and neurodegenerative disorders22,23,24,25,26. Condensate formation is often driven by transient, selective, and multivalent protein-protein, protein-nucleic acid, or nucleic acid-nucleic acid interactions27. Under certain conditions, these interactions can lead to liquid-liquid phase separation (LLPS), a density transition that locally enriches specific biomolecules in membraneless droplets. Such multivalent interactions are often mediated by the intrinsically disordered regions (IDRs) of proteins1,28,29. Biophysical characterization of these interactions at the molecular level is critical to our understanding of numerous healthy and aberrant cellular functions, given the pervasiveness of condensates across them. Although techniques based on confocal fluorescence microscopy, e.g., fluorescence recovery after photobleaching (FRAP)30,31,32, have been widely used to qualitatively show that the molecular exchanges between condensates and the surrounding cellular environment are dynamic, quantifying the interaction dynamics of specific biomolecules within condensates is generally not possible using conventional confocal microscopy or single-molecule microscopy without specialized data analysis methods. The single-particle tracking (SPT) technique described in this protocol is based on live-cell single-molecule microscopy33 and provides a uniquely powerful tool to quantify the interaction dynamics between specific proteins within condensates. The readout of SPT for such measurement is the mean residence time of a protein of interest in the condensates.
The protocol can be broken down into two parts – data acquisition and data analysis. The first step of imaging data acquisition is to express in cells a protein of interest that is fused to a HaloTag34. This enables labeling of the protein of interest with two fluorophores, where a majority of the protein molecules are to be labeled with a non-photoactivatable fluorophore (e.g., JFX549 Halo ligand35) and a small fraction of them are to be labeled with a spectrally distinct, photoactivatable fluorophore (e.g., PA-JF646 Halo ligand36). This allows for the simultaneous acquisition of all condensate locations in the cell and the acquisition of single-molecule movies of the protein of interest binding and unbinding to the condensates. Meanwhile, the same type of cells are modified to stably express Halo-tagged H2B, a histone that is largely immobile on chromatin. The cells are then stained with the PA-JF646 Halo ligand to enable single-molecule imaging of H2B. As will be discussed in detail below, this experiment accounts for the contribution of photobleaching to enable precise quantification of the interaction dynamics of the protein of interest. Cells for imaging experiments must then be cultured on clean coverslips, stained with HaloTag ligand(s), and assembled into a live-cell imaging chamber. From there, the sample is imaged under highly inclined and laminated optical sheet (HILO) illumination on a total internal reflection fluorescence (TIRF) microscope capable of two-channel imaging and single-molecule detection. The emission is then split onto two cameras, one tracking condensate positions and one tracking single molecules. Acquisition is performed with a long integration time (on the order of hundreds of ms) to blur out freely-diffusing proteins and only capture proteins that are less mobile due to binding to stable structures in the cell37.
The first step of data analysis is using an established single-particle tracking (SPT) algorithm38,39 to localize individual protein molecules in each frame of the movie and assemble the localizations into a trajectory for each molecule over its detectable lifetime. The trajectories are then sorted into those representing molecules inside and those representing molecules outside the condensates by comparing the localizations of the molecules throughout their trajectories to the localizations of all the condensates at the corresponding times1.
Next, a survival curve (1 – CDF) is generated using the lengths of all the in-condensate trajectories. The apparent mean residence time of the molecules is then extracted by fitting the survival curve to the following two-component exponential model of protein binding,
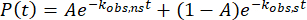 ,
,
with A as the fraction of molecules non-specifically bound and with kobs,ns and kobs,s as the observed dissociation rates of the non-specifically bound and specifically bound molecules, respectively. Only kobs,s is considered from here onward. The dynamics of both protein dissociation, ktrue,s, and photobleaching of the fluorophore, kpb, contribute to kobs,s as
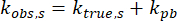 ;
;
thus, to isolate the effects of protein dissociation, the specific dissociation rate of H2B-Halo in the cell line mentioned prior is measured.
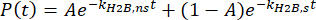
H2B is a protein that is stably integrated into chromatin and that experiences minimal dissociation in the time scale of a single-molecule movie acquisition37. Its specific dissociation rate is then equal to the photobleaching rate of the PA-JF646 Halo ligand, or
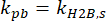 .
.
The mean in-condensate residence time of the protein of interest, 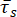 , is then
, is then
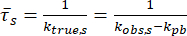 .
.
Representative results from Irgen-Gioro et al.40 are shown, where this protocol was applied to demonstrate that fusing an oligomerization domain to IDR results in longer residence times of the IDR in its condensates. This result suggests that the added oligomerization domain stabilizes the homotypic interactions of the IDR that drives LLPS. In principle, the same method with slightly modified protocols can be applied to characterize the homotypic or heterotypic interactions of any protein that participates in the formation of any types of condensates.
