An architecture for modular intracellular protein detection
As shown in Figure 1, the device is composed of: 1) intrabody 1 connected to the membrane-tethered fluorescent marker (mKate) and TEVp cleavage site (TCS), followed by a transcription activator GAL4VP16 (TF); 2) intrabody 2 fused to TEV protease (TEVp), free in the cytosol; 3) a synthetic promoter responsive to GAL4VP16, driving the expression of a reporter gene. The modularity is guaranteed by intrabodies that can be changed in a plug-and-play manner to target any desired protein. Also, the output can be chosen depending on the desired application. A critical aspect in order to optimize sensing-actuation performance is to avoid background activation of the system (in absence of the protein of interest). We thus tuned several features of the device (Table 1).
HCV, Huntingtin and HIV sensor-actuator devices
The first device was built to recognize NS3 protein, expressed from the epatitis C virus HCV29. The study of this sensor set the rules for the following ones. In specific, we co-transfected fusion protein 1 variants of the NS3-responsive devices along with constitutively expressed (hEF1α) TEVp-scFv162 in HEK293FT cells. We observed NS3-independent output activation both with the TCS(S) and TCS(L) (Figure 2a), indicating that the system requires fine regulation of TEVp activity. To achieve input specific signal activation, we modulated TEVp expression transcriptionally and post-translationally. The first was mediated by a doxycycline (dox) responsive promoter (pTET), whereas the second was obtained by adding to TEVp a degradation domain tag (DD degron) regulated by the small molecule shield. The device was able to specifically induce gene expression following NS3 detection, in the absence of dox and shield when the system included the TCS(L) (Figure 2b). These results indicate that minimal TEVp concentration is required for efficient devices. On the contrary maximal activation of TEVp by dox and shield in the TCS(L) configuration, or the TCS(S) exhibits high background (Figure 2b, c). Altogether, the results demonstrate that the simple leakiness of the promoter (absence of doxycycline or the activator rtTA3) enables the best ON/OFF condition, highlighting the high sensitivity of the TEVp actuator.
Based on the results we designed further NS3 device variants by changing parameters previously mentioned (Table 1) and demonstrated up to 24 EYFP fold induction (Figure 2d,f), and specific cell killing (data in20). Further, we fused the NS3 to a blue fluorescent protein and demonstrated colocalization with fusion protein 1, indicating the interaction between the component of the system (Figure 2e).
Nef sensor-actuator device
Differently from the previous example, the sensing module of Nef device is composed of a single domain antibody (sdAb19)31, and SH3 domain from p59fyn protein tyrosine kinase that is highly expressed in T lymphocytes and implicated in antigen induced T-cell activation34. Since Nef is involved in HLA-I downmodulation, we hypothesized that by sequestering Nef, the device could impair its pathological activity (depicted in Figure 3).
We first tested the Nef-responsive device in 293FT HEK and Jurkat T cells. The sensor exhibited up to 5-fold induction of EYFP in the presence of the target protein (Figure 4a). Different from the NS3 case, here the activation of TEVp by dox (D100) was required to obtain a functional device, whereas LD0 demonstrated lower background in the absence of the target protein. Moreover, we observed EYFP expression in Jurkat T cells infected with X4 LAI strain (Figure 4b). We also demonstrate that the device impairs Nef-mediated downregulation of HLA-I (Figure 4c). Finally, we rewired the output to induce XCL-1, a chemokine normally secreted by CD8+ T cells to induce a localized CTL response, demonstrating the potential towards selective activation of the immunomodulator (Figure 4d).
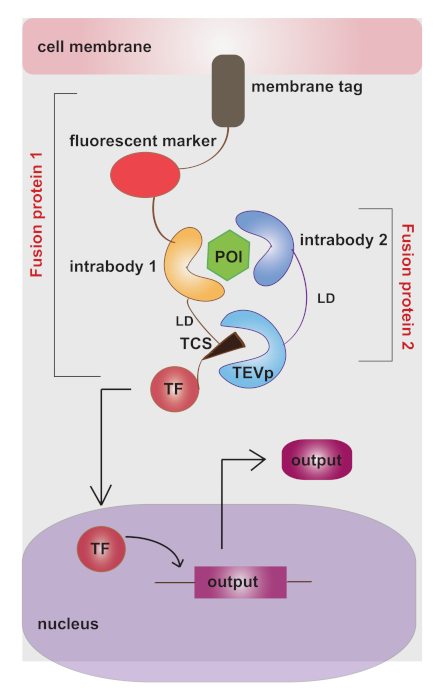
Figure 1: Architecture of the sensor-actuator device for intracellular protein detection. The device is composed of intrabodies (or small peptides) to recognize with high specificity the protein of interest (POI). Interaction of intrabodies with POI brings TEVp in the proximity of the cleavage site (TCS). The membrane-tethered transcription factor (TF) fused to TCS translocates to the nucleus upon TEVp cleavage, activating programmed output. The figure has been modified from20. Please click here to view a larger version of this figure.
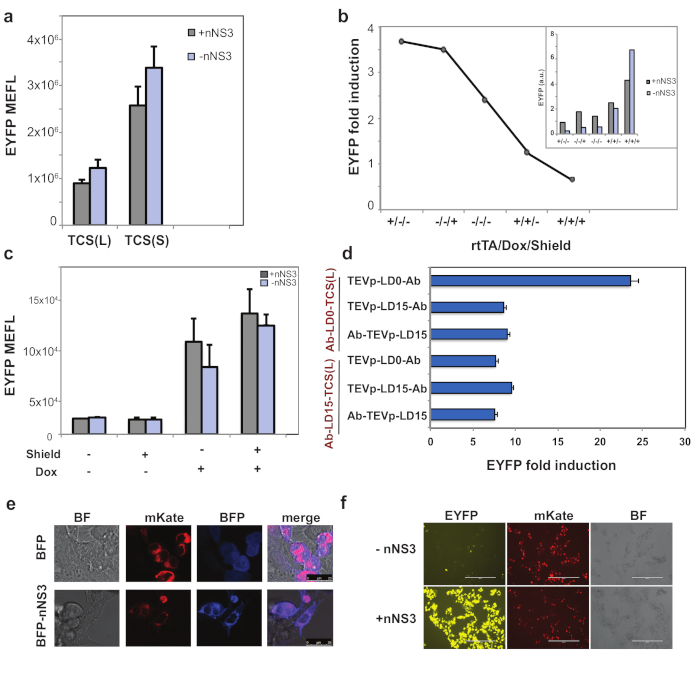
Figure 2: Test of the NS3 device. (a) TCS(S) and (L) variants of NS3 device were tested in HEK293FT cells in presence or absence on NS3. TEVp is driven by a constitutive promoter. Data show that the system is not sensitive to the presence of NS3 due to an overactivation of TEVp that leads to non-specific cleavage of the TCS. (b-c) NS3 induced EYFP reporter activation with TCS(L) but not TCS(S) in absence of Dox and Shield. (d) Test of the variants of NS3 sensor in HEK293FT cells show up to 24x induction of reporter gene when the HVC protein was detected. Data shows fold induction and standard deviation. (e) Co-localization in HEK293FT cells of NS3 fused to a BFP (BFP-nNS3) and fusion protein 1 which include the membrane tethered mKate. As control we used a BFP alone, that shows diffuse cellular localization. Confocal images (scale bar = 25 μm) (f) fluorescent reporter induction as result of NS3 detection in HEK293FT cells. BF: bright field; mKate: fluorescent marker included in fusion protein 1; EYFP: output of the device. (scale bar = 200 μm). The figure has been modified from20. Please click here to view a larger version of this figure.
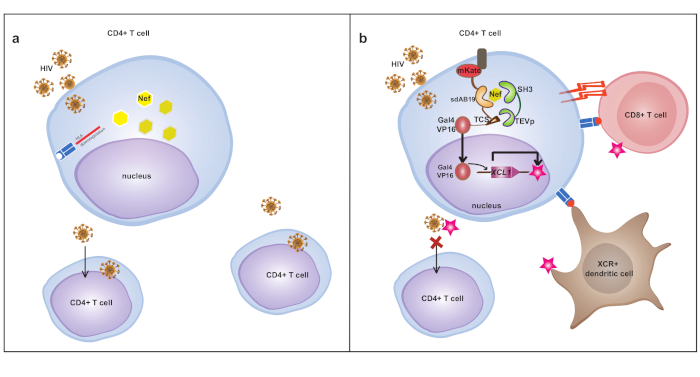
Figure 3: Model of Nef-responsive device. (a) In the absence of the engineered device, HIV escapes the immune response via Nef-mediated HLA downregulation. (b) Nef is detected and sequestered by the sdAb19 intrabody and SH3 domain to activate XCL1 expression. XCL1 stimulates anti-HIV immune response attracting dendritic cells and cytotoxic lymphocytes. In addition, Nef sequestration impairs HLA downmodulation such that CD4+T cells infected by HIV are more responsive to immune system attack. The figure has been modified from20. Please click here to view a larger version of this figure.
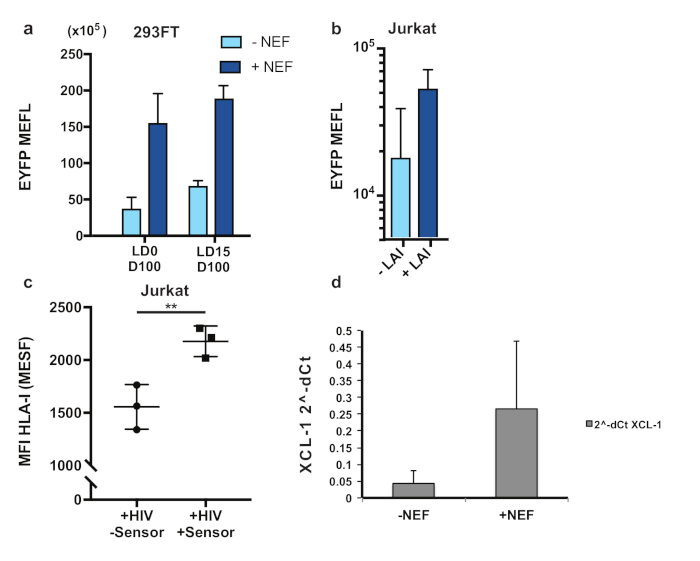
Figure 4: Nef sensor-actuator device detects HIV infection and impair HLA downmodulation. (a) HEK293FT cells induce output activation in the presence of Nef, at intermediate concentration of Dox. (b) Jurkat T cells infected with HIV-LAI strain show reporter activation. (c) Nef device interferes with HLA downmodulation as compared to HLA levels in absence of sensor. (d) XCL-1 immunomodulator is triggered by the device upon Nef detection. The figure has been modified from20. Please click here to view a larger version of this figure.
| 1. Regulation of TEVp expression | |||||
| Transcriptional | Constitutive (hEF1a) | ||||
| Inducible (pTET) | |||||
| Post-translational | Degradation Domain tag (DD degron) + Shield | ||||
| 2. TEV Cleavage Site (TCS) | |||||
| Aa substitutions in P1 | Affinity | ||||
| TCS-S | Serine | High | |||
| TCS-L | Leucine | Low | |||
| 3. Position of TEVp | |||||
| N-terminus of intrabodies | |||||
| C-terminus of intrabodies | |||||
| 4. Glycine-Serine (G4S) Linker Domains (LD) | |||||
| LD0 | 0 Amino acids | ||||
| LD10 | 10 Amino acids | ||||
| LD15 | 15 Amino acids | ||||
| 5. Intrabodies and analogues | |||||
| NS3 | HTT | Tat | Nef | ||
| Intrabody 1 | scFv35 | Happ1 | scFv2 | sdAb19 | |
| Intrabody 2 | scFv162 | VL12.3 | scFv3 | SH3* | |
Table 1: Components for modular device design. Construction of efficient modular devices requires several elements, which can be adjusted to improve the system sensitivity, and specificity. TEVp activity can be adjusted in order to achieve the best signal-to-noise ratio in the system: (1) via transcriptional modulation of TEVp expression using a constitutive (hEF1α) or Doxycycline inducible promoter (pTET), or via posttranslational modulation, using a degradation domain tag (DD) controlled by the small molecule Shield; (2) via high or low affinity TEV cleavage sites (TCS) that include a serine (TCS-S) or a leucine (TCS-L) in P1 position; (3) via repositioning of TEVp to the N or C terminus of the intrabody. To take into account the intrabodies-protein interaction and TEVp accessibility to TCS we (4) included Glycine-Serine (G4S) Linker Domains (LD) of variable amino acid length between intrabody and TCS and between TEVp and intrabody. (5) Selection of intrabodies or their analogues to the target protein. Of note, *SH3 of the Nef-responsive device is not an intrabody but a domain of p59fyn protein tyrosine kinase that bind a proline-rich motif in Nef32. These components were assembled in a plug-and-play fashion to build a modular system towards optimal input-output devices.
| Parameters of the protein sensor-actuator devices | |
| TEVp affinity to the cleavage site | |
| TCS (S) | ENLYFQ S |
| TCS (L) | ENLYFQ L |
| TEVp activity modulation by small molecules | |
| Transcriptional | Doxycycline |
| Translational | Shield |
| TEVp access to the TCS | |
| Fusion protein design | N- or C- terminus fusion o TEVp to the intrabody |
| Protein flexibility | |
| Flexible Linker (LD) | no linker |
| 10 or 15Aa (G4S)3 |
Table 2: Modules of the best NS3 device. Best performing sensory-actuator device is reported for NS3. Precisely, we specify the length of LD domain (LD0, LD15), affinity of the protease binding site TCS (S, L), type of TEVp regulation (hEF1α or pTET, or DD degron).
| Principal engineered modules | ||
| Components | Short name | |
| Nef | Protein of interest | Nef |
| Fusion protein 1 | phEF1a-sdAb19-LD0-TCS(L) | |
| Fusion protein 2 | pTET-TEVp-LD0-SH3 | |
| Output (cytokine) | UAS-XCL1 | |
| Output (fluorescence) | UAS-EYFP | |
Table 3: Modules of the best Nef device. Best performing sensory-actuator device is reported for Nef. Precisely, we specify the length of LD domain (LD0, LD15), affinity of the protease binding site TCS (S, L), type of TEVp regulation (hEF1α or pTET).
