Stimulating AdMSCs with TNFα or interferon-gamma (IFNγ) for 24 h induces changes in the expression of anti-inflammatory molecules and growth factors. Treating AdMSCs with TNFα or interferon-gamma (IFNγ) increases TNF-stimulated gene 6 (TSG-6) mRNA, whereas TNFα, but not IFNγ, causes an increase in transforming growth factor beta-1 (TGFβ-1) mRNA. Stimulation with TNFα or IFNγ induces an increase in vascular endothelial growth factor (VEGF)mRNA, but no change in the expression of fibroblast growth factor (FGF) mRNA (Figure 1A). These data suggest that AdMSCs likewise respond to factors in prion-infected brain homogenate, likely a milieu of cytokines and damage-associated molecular patterns (DAMPs) 18,19,20. We show that culturing AdMSCs for 24 h in media containing 0.1% normal brain homogenate (NBH) or Rocky Mountain Laboratories (RML) strain mouse-adapted scrapie brain homogenate increases expression of TSG-6 mRNA. Treatment with RML, but not NBH, increased mRNA for TGFβ-1, VEGF, and FGF (Figure 1B).
To assess the role AdMSCs play in decreasing inflammation in both microglia and astrocytes, primary mixed glia were infected with 0.1% NBH or 22L mouse-adapted scrapie. At 7 days post infection, a co-culture system was established by adding AdMSCs to inserts and co-cultures for 7 days (Figure 2A). Analysis of the mixed glia demonstrated a significant increase in mRNA for the inflammatory cytokines CCL2, CCL5, and IL1β, and the astrocyte marker S100β, in cells infected with 22L compared to NBH. No significant changes were seen in mRNA expression for TNFα. After co-culturing for 7 days with AdMSCs, a decrease in CCL2, CCL5, and IL1β mRNA was seen in both NBH-treated and 22L-infected cells, but no significant changes were seen in TNFα mRNA expression. Co-culturing with AdMSCs also decreases the pan-astrocyte marker S100β and the reactive astrocyte marker C3 in both NBH and 22L-treated glia (Figure 2B).
The BV2 microglia cell line was treated with NBH or RML to assess changes specific to microglia after culturing with AdMSCs. Because stimulating AdMSCs with TNFα causes an increase in the production of the anti-inflammatory gene, as described above, AdMSCs were stimulated for 24 h prior to co-culturing with BV2 cells. At 6 days post prion exposure, BV2 cells were co-cultured with AdMSCs (Figure 2C). RNA was isolated after 24 h. Although we do not show any differences in inflammatory markers between NBH and RML-treated BV2 cells, we show that AdMSCs drastically decrease the transcription of pro-inflammatory markers in BV2 cells. A significant decrease was seen in the inflammatory cytokines IL1β, IL-6, and TNFα and the complement protein C1qa for both NBH-treated and RML-treated BV2 cells. Co-culturing with AdMSCs decreased the M1 microglial gene CD-16 and increased mRNA for Arg-1, a marker for M2 microglia, in both NBH-treated and RML-treated BV2 cells (Figure 2D).
Not only does stimulating MSCs increase the production of anti-inflammatory molecules and growth factors, but it also improves their ability to migrate to areas of inflammation 13,21. Here, we demonstrate an in vitro assay to assess AdMSC migration toward the prion-infected brain. AdMSCs can be stimulated with TNFα for 24 h, then serum-starved for 4 h, then added to inserts above wells containing media only, or media with 1% NBH or RML (Figure 3A). After 24 h, some cells migrate toward media only and NBH-containing media, but there is increased migration of cells toward RML-containing media (Figure 3B,C).
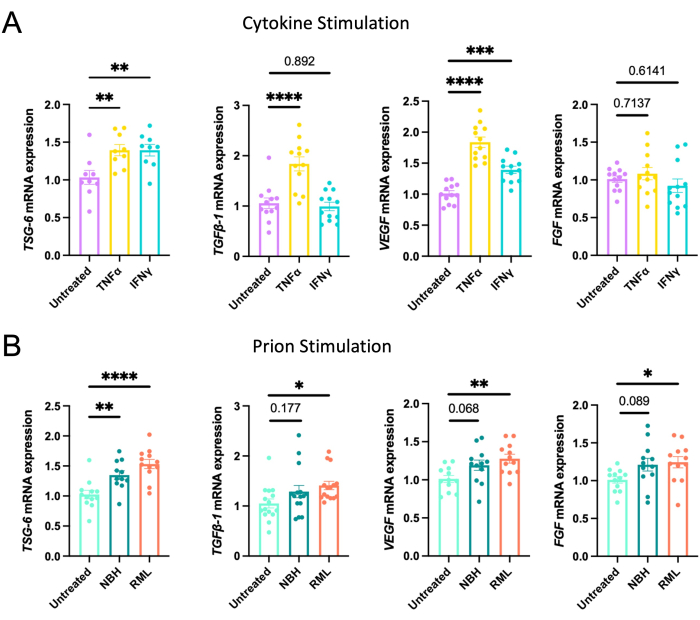
Figure 1: Induction of the expression of anti-inflammatory genes and growth factors after stimulating AdMSCs with cytokines or prion-infected brain homogenate. AdMSCs at passage 3 were stimulated for 24 h with (A) 10 ng/mL TNFα, which induced the expression of mRNA for TSG-6, TGFβ-1, and VEGF, or with 200 ng/mL IFN-α for 24 h induced the expression of TSG-6 and VEGF. (B) Stimulation for 24 h with 0.1% NBH induced expression of TSG-6 mRNA. Stimulation with 0.1% RML prion-infected brain homogenate induced expression of mRNA for TSG-6, TGFβ-1, VEGF, and FGF. One-way ANOVA with post-hoc Tukey's test, *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, error bars = SEM. Combined data from four separate experiments, each with three technical replicates. This figure is adapted from Hay et al.22 Abbreviations: AdMSCs = adipose-derived mesenchymal stromal cells; TNFα = tumor necrosis factor alfa; IFN-α = interferon-gamma; TSG-6 = TNF-stimulated gene 6; TGFβ-1 = transforming growth factor beta-1; VEGF = vascular endothelial growth factor; NBH = normal brain homogenate; RML = Rocky Mountain Laboratory prion strain. Please click here to view a larger version of this figure.
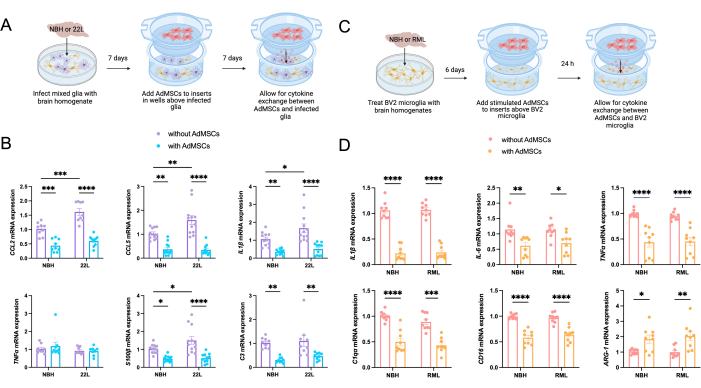
Figure 2: Decrease in markers of inflammation in prion-treated primary mixed glia and BV2 microglia following co-culture with AdMSCs. (A) Primary mixed glia were infected with 0.1% NBH or 22L prion-infected brain homogenate for 72 h, thoroughly washed, then cultured in normal media for 96 additional h. AdMSCs were added to inserts above the corresponding wells. Glia-AdMSC cocultures were incubated for an additional 7 days, for a total of 14 days from the initial infection. (B) Glia infected with 22L showed increased expression of mRNA for the inflammatory markers CCL2, CCL5, and IL1β, in addition to the astrocyte marker S100β compared to those treated with NBH. Co-culturing with AdMSCs decreased expression of the inflammatory cytokines CCL2, CCL5, and IL1β, but not TNFα, for both NBH and 22L-infected cells, as well as decreasing expression of S100β and C3, which together are markers of reactive astrocytes. (C) BV2 microglia were treated with 0.1% NBH or RML prion-infected brain homogenate for 72 h, thoroughly washed, then cultured in normal media for 72 additional h. AdMSCs were stimulated with 10 ng/mL TNFα for 24 h, then added to inserts above corresponding wells. BV2-AdMSC co-cultures were incubated for 24 h, for a total of 7 days from initial brain homogenate treatments. (D) BV2s co-cultured with AdMSCs showed a decrease in mRNA for the inflammatory markers IL1β,IL-6, and TNFα, and the complement protein C1qa. Additionally, AdMSCs decreased the M1 microglia gene CD-16 and increased the M2 microglia marker Arg-1. Two-way ANOVA with post-hoc Tukey's test, *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, error bars = SEM. Combined data from three separate experiments, each with three technical replicates. This figure is adapted from Hay et al.14. Abbreviations: AdMSCs = adipose-derived mesenchymal stromal cells; IL = interleukin; TNFα = tumor necrosis factor alfa; NBH = normal brain homogenate; 22L = 22L mouse-adapted scrapie prion strain. Please click here to view a larger version of this figure.
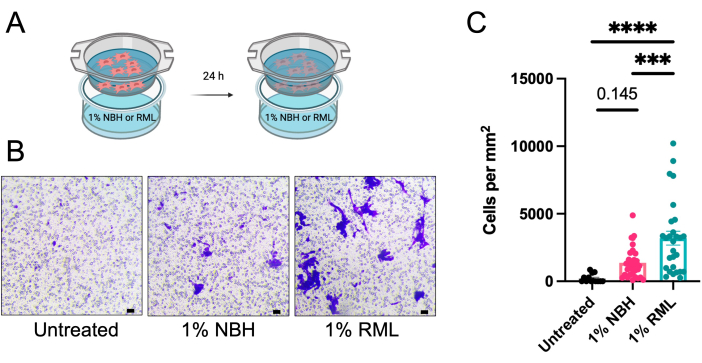
Figure 3: AdMSCs migration towards prion-infected brain homogenate in an in vitro model. AdMSCs were stimulated for 24 h with 10 ng/mL TNFα, then serum-starved for 4 h. (A) They were plated in inserts with a pore size of 8 µm above media alone, or media containing 1% NBH, or 1% RML prion-infected brain homogenate, and incubated for 24 h. (B) AdMSCs were stained with crystal violet solution and cells that had migrated to the bottom side of the insert were imaged and counted manually. (C) Migration of AdMSCs toward NBH-containing media is not significantly more than toward untreated media. However, AdMSCs show significant migration toward media containing 1% RML. Data are a combination of three biological replicates each with three technical replicates, with four fields of vision for each technical replicate. One-way ANOVA with post-hoc Tukey's test, **p < 0.001, ****p < 0.0001, error bars = SEM. Scale bars = 10 µm (B). This figure is adapted from Hay et al.14. Abbreviations: AdMSCs = adipose-derived mesenchymal stromal cells; TNFα = tumor necrosis factor alfa; NBH = normal brain homogenate; RML = Rocky Mountain Laboratory mouse-adapted scrapie prion strain. Please click here to view a larger version of this figure.
Table 1: Murine primer sequences for reverse transcriptase quantitative PCR. Please click here to download this Table.
