Начиная с первоначального "мозаики жидкости" модели Зингера и Николсона, картина сотовой мембране постоянно обновляется в течение последних десятилетий, чтобы включить возрастающую роль цитоскелета и липидных доменов 1,2.
Первые наблюдения были получены методом флуоресцентной восстановления после фотообесцвечивания (FRAP) открытие, что значительная часть мембранных белков неподвижен 3-5. Эти пионерские исследования, хотя и очень информативным, пострадали от относительно низкого разрешения в пространстве (микрон) и время (в секундах) FRAP установок. Кроме того, будучи ансамбля измерение усреднения, FRAP не хватает в предоставлении информации о поведении одной молекулы.
В этом контексте, возможность специально маркировать одну молекулу с очень яркими тегов (позволяющих изучение процесса диффузии одной молекулы в то время) был очень успешным. В частности, нажаввременное разрешение на Одноместный Отслеживание частиц (SPT) подхода к шкале времени микросекунд, Кусуми и др. получили доступ к неизвестных особенностей динамики липидных и белковых что значительный вклад в признание роли актина основе мембранной скелета в мембранной физиологии 6 , 7. Эти данные генерируются так называемый "пикет и забор 'модель, в которой липидов и белков диффузии регулируется актина на основе скелета. Тем не менее, для того чтобы иметь доступ к огромному количеству информации, представленной ППП многих экспериментальных вопросов должны решаться. В частности, процедура маркировки, как правило, состоит из многих шагов, таких как производства, очистки и введения меченых видов в системе. Кроме того, большие наклейки, как квантовых точек или металлических наночастиц, часто требуется, чтобы достичь доли миллисекунды, временные рамки и сшивание молекул-мишеней по этикетке не можно было бы избежать во многих случаях. Наконец, многие траекториикоторые должны быть записаны в соответствии статистические критерии, и одновременно низкой плотности этикетки необходимо, чтобы отслеживать.
По сравнению с SPT, корреляционной спектроскопии флуоресценции (ФТС), преодолевая многие из этих недостатков, представляет собой очень перспективный подход к изучению молекулярной динамики. На самом деле, ФТС работает хорошо и с тусклыми и плотных этикеток, что позволяет изучать динамику флуоресцентных белков с метками молекул в временно трансфицированных клеток. Кроме того, это позволяет достичь высоких статистики в ограниченный период времени. Наконец, несмотря на "высокий" плотности этикеток FCS обеспечивает Одиночные молекулы информацию. Благодаря всем этим свойствам, ФТС представляет собой очень простой подход и широко применяется для изучения динамики липидов и белков как в модельных мембран и живых клеток 8-10. Многие различные подходы были предложены для повышения способности ФТС раскрыть детали молекулярной диффузии. Например, это было SHсамостоятельно, что, выполняя FCS на разного размера смотровых площадок можно определить "закону диффузии FCS" просветительских скрытые черты молекулярного движения 11,12. Помимо того, что изменились в размере, фокусное район был также дублируется 13, перемещается в пространстве по линиям 14-20 или сопряженных с быстрыми камерами 21,22. Использование этих "пространственно-временной" корреляцию подходы, соответствующие биологические параметры нескольких мембранных компонентов были количественно описаны на обеих модельных мембран и фактических биологических, получая таким образом понимание мембраны пространственной организации.
Тем не менее, во всех FRAP и приложений ФТС описанные до сих пор размер фокальной области представляет собой предел в пространственным разрешением, что не могут быть преодолены. Несколько методов визуализации супер-разрешения были недавно разработаны, чтобы обойти это ограничение. Некоторые из них основаны на точность локализации, такие как стохастической оптической микроскопии реконструкции (STORM) <вир> 23,24, локализация фотоактивация микроскопии (PALM) 25, флуоресценции PALM (FPALM) 26, и отслеживание PALM одночастичное (sptPALM) 27: относительно большое количество фотонов, необходимых на каждом снимке, однако, ограничивает временное разрешение эти методы, по крайней мере несколько миллисекунд, тем самым, препятствуя их применимость в естественных условиях.
В отличие от этого, перспективной альтернативой для визуализации суперразрешением были открыты пространственно модуляции флуоресцентное излучение с стимулированных методов истощения выбросов (STED или обратимых насыщающихся оптических переходов флуоресценции (RESOLFT)) 28,29. Эти подходы сочетают формирование объема наблюдательной скважине ниже дифракционного предела с возможностью использования быстрого сканирования микроскопы и системы обнаружения. В сочетании с анализом флуоресценции флуктуации, STED микроскопии позволило непосредственно исследовать наноуровне пространственно-временные динамику липидов и рroteins в живых клеточных мембран 30,31.
Те же физические величины STED основе микроскопии можно получить с помощью модифицированной пространственно-временной корреляции изображения спектроскопии (Stics 32,33) метод, который подходит для изучения динамики флуоресцентно-меченых мембранных белков и / или липидов в живых клетках и коммерческой микроскопом. Экспериментальный протокол, представленный здесь состоит несколько шагов. Первый требует быстрого получения изображения области интереса на мембране. Затем полученный Стек изображений используется для вычисления средних пространственно-временные корреляционные функции. Установив серию корреляционные функции, молекулярная 'диффузия закон' могут быть получены непосредственно из изображений в виде кажущейся диффузии (D приложения) – против усредненный участка смещения. Этот участок критически зависит от окружающей среды изучены молекулами и позволяет признавая непосредственно фактические режимы диффузиилипидной / интересующего белка.
Более подробно, как было показано ранее 34, пространственно-временное авто-корреляционная функция приобретенной серии изображений критически зависит от динамики молекул, движущихся в серии собраны изображения (обратите внимание, что те же самые рассуждения могут быть применены в приобретении линии где лишь одна в пространстве считается). В частности, мы определяем функцию корреляции, так:
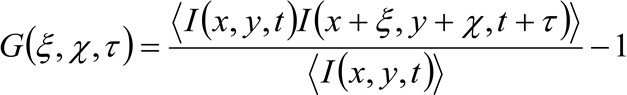 (1)
(1)
где 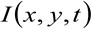 представляет измеренное интенсивности флуоресценции в положении х, у и в момент времени т,
представляет измеренное интенсивности флуоресценции в положении х, у и в момент времени т, ![]() и
и ![]() представляет собой расстояние по оси х иу направления соответственно,
представляет собой расстояние по оси х иу направления соответственно, ![]() представляет собой временной лаг, и
представляет собой временной лаг, и ![]() представляет собой среднее. Эта функция может быть выражена как:
представляет собой среднее. Эта функция может быть выражена как:
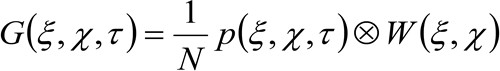 (2)
(2)
где 'N' представляет собой среднее число молекул в зоне наблюдения, ![]() представляет операцию свертки в пространстве, и
представляет операцию свертки в пространстве, и 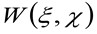 представляет автокорреляции инструментальной талии. Этот последний может быть интерпретирована как мера того, насколько фотоны одной излучателя разбросаны в пространстве за счет оптического / Recording Setup (так называемый функция рассеяния точки, PSF, генРалли хорошо аппроксимируется функцией Гаусса). И, наконец,
представляет автокорреляции инструментальной талии. Этот последний может быть интерпретирована как мера того, насколько фотоны одной излучателя разбросаны в пространстве за счет оптического / Recording Setup (так называемый функция рассеяния точки, PSF, генРалли хорошо аппроксимируется функцией Гаусса). И, наконец, 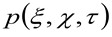 представляет собой вероятность найти частицу на расстоянии
представляет собой вероятность найти частицу на расстоянии ![]() и
и ![]() после задержки времени
после задержки времени ![]() . Если учесть, диффузным динамику, в котором частицы движутся беспорядочно во всех направлениях и чистые потоки не присутствуют, эта функция также хорошо аппроксимируется функцией Гаусса, где дисперсия может быть идентифицирован как средний квадрат смещения (MSD) движущейся частицы . Таким образом, талия корреляционной функции (также упоминается как
. Если учесть, диффузным динамику, в котором частицы движутся беспорядочно во всех направлениях и чистые потоки не присутствуют, эта функция также хорошо аппроксимируется функцией Гаусса, где дисперсия может быть идентифицирован как средний квадрат смещения (MSD) движущейся частицы . Таким образом, талия корреляционной функции (также упоминается как ![]() ), Может быть определен как сумма опорно частиц и инструментальной талии и может быть измерена с помощью гауссовой подгонкитин корреляционной функции для каждого времени задержки. Измеренное я МСД может быть использована для расчета кажущейся диффузии подвижных молекул
), Может быть определен как сумма опорно частиц и инструментальной талии и может быть измерена с помощью гауссовой подгонкитин корреляционной функции для каждого времени задержки. Измеренное я МСД может быть использована для расчета кажущейся диффузии подвижных молекул ![]() и среднее смещение
и среднее смещение ![]() как:
как:
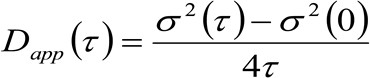 (3)
(3)
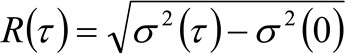 (4)
(4)
Несколько соображений по экспериментальной установки можно вести читателя на протяжении следующих разделах. Для того, чтобы избирательно возбуждать флуорофоры на базальной мембране живой клетки мы будем использовать полное внутреннее отражение (TIR) освещение, используя коммерческий флуоресценции МДП (TIRF) микроскоп (подробности можно найти в разделе материалов). Кроме того, для того, чтобы собрать йэлектронной флуоресценции мы будем использовать большом увеличении цели (100X NA 1,47 высокой числовой апертурой требуется для освещения TIRF) и EMCCD камеры (физический размер пикселя на чип 16 мкм). Чтобы достичь размер пикселя 100 нм мы применяем дополнительную увеличением объектива фактор 1.6. Как будет показано ниже, временное разрешение ниже 1 мс необходимо будет правильно описывать динамику быстрых липидов мембран ниже 100 нм. Для того, чтобы достичь этой временное разрешение нам нужно выбрать область процентной (ROI) меньше, чем всего чипа камеры (512 х 512). Таким образом, камера будет читать уменьшенное количество линий увеличения временного разрешения. Тем не менее, в этом считывания режима сроки будет ограничено на время, необходимое для сдвига зарядов от воздействия на чип считывания на камеру и, как правило, в порядке миллисекундах для 512 х 512 пикселей EMCCD. Чтобы побить этот предел, новая технология позволяет сдвига ROI-линии только вместо целого кадра, жIth практической эффективного снижения открытой размеров кристалла (так называемый режим Обрезанные Датчик в нашей EMCCD). Для этой конфигурации, чтобы быть эффективным, чип за пределами ROI должны быть покрыты пару прорезей, установленных в оптическом пути. Благодаря этой установке временным разрешением до 10 -4 секунд может быть достигнуто. Пожалуйста, обратите внимание, однако, что этот подход может быть связан с различными экспериментальных установок, как описано в разделе «обсуждение».
Демонстрация метода будут представлены в живых клетках, с использованием как ATTO488 помечены 1-пальмитоил-2-гидрокси-Sn глицеро-3-фосфоэтаноламин (ATTO488-PPE) и GFP-меченых вариант рецептора трансферрина (GFP- TfR). В случае ATTO488-PPE этот подход может успешно восстанавливать почти постоянное D приложение в зависимости от среднего смещения, указывающего в основном свободную диффузию, как сообщалось ранее 30,35. В отличие от этого, TFR-GFP показывает убывающую D <suб> приложение в зависимости от среднего водоизмещения, предлагая частично ограниченном диффузию 6. Более того, в последнем случае можно дать количественную оценку локального коэффициента диффузии и среднюю области удержания в течение многих мкм на плоскости мембраны.
