Med utgångspunkt från den ursprungliga "vätske mosaik" modell av Singer och Nicolson, bilden av cellplasmamembranet har kontinuerligt uppdaterats under de senaste decennierna för att inkludera den framväxande roll cytoskeleton och lipid domänerna 1,2.
De första observationerna erhölls genom fluorescerande återhämtning efter fotoblekning (FRAP) avtäckningen att en betydande fraktion av membranproteiner är orörlig 3-5. Dessa banbrytande studier, även om mycket informativt, led av den relativt dåliga upplösningen i rymden (mikron) och tid (sekunder) i FRAP inställningar. Dessutom är en ensemble medelvärdesmätning, FRAP saknar i att ge information om enda molekyl beteende.
I detta sammanhang är det möjligt att specifikt märka en enda molekyl med mycket ljusa taggar (möjliggör studiet av diffusionsprocessen en molekyl i taget) har varit mycket framgångsrikt. Speciellt, genom att trycka påtidsupplösning av den inre Particle Tracking (SPT) inställning till mikrosekunder tidsskalan, Kusumi, et al. fått tillgång till okända funktioner i lipid och proteindynamik som kraftigt bidragit till erkännandet av den roll aktin baserade membranskelett i membranfysiologi 6 , 7. Dessa fynd genererade den så kallade "strejkvakt och staket" modell, där lipid och protein diffusion regleras av aktin baserade skelettet. Men för att få tillgång till den enorma mängd information som SPT många experimentella frågor måste lösas. Speciellt är märkningsförfarandet vanligen består av många steg som produktion, rening och införande av de märkta arter i systemet. Dessutom stora etiketter, som kvantprickar eller metallnanopartiklar, ofta krävs för att nå under millisekund tid och tvärbindningen av målmolekyler med etiketten kunde inte undvikas i många fall. Slutligen, många banormåste registreras för att passa statistiska kriterier och samtidigt en låg densitet av etiketten krävs för att möjliggöra spårning.
Jämfört med SPT, fluorescens-korrelationsspektroskopi (FCS), övervinna många av dessa nackdelar, är ett mycket lovande tillvägagångssätt för att studera molekylär dynamik. I själva verket fungerar FCS bra även med dim och täta etiketter, som gör det möjligt att studera dynamiken i fluorescerande protein-märkta molekyler i gående transfekterade celler. Dessutom möjliggör det att nå höga statistik i en begränsad tid. Slutligen, trots den "höga" densitet av etiketter FCS erbjuder enstaka molekyler information. Tack vare alla dessa egenskaper, FCS är en mycket enkel metod och har tillämpats i stor utsträckning för att studera lipid och protein dynamik både i modellmembran och i levande-celler 8-10. Många olika metoder har föreslagits för att öka förmågan hos FCS att avslöja detaljerna i molekylär diffusion. Till exempel var det shegna som genom att utföra FCS på olika stora observationsområdena kan man definiera en "FCS diffusion lag" upplysande dolda funktioner i molekylär rörelse 11,12. Förutom att vara varierade i storlek, var det prioriterade området också dupliceras 13, flyttade i rymden längs linjerna 14-20 eller konjugerade med snabba kameror 21,22. Med hjälp av dessa "tid och rum" korrelation tillvägagångssätt, var relevanta biologiska parametrar för flera membrankomponenter kvantitativt beskrivas både modellmembran och faktiska biologiska sådana, vilket således ger inblick i membran rumslig organisation.
Men i alla FRAP och FCS-program hittills beskrivna storleken på det prioriterade området utgör en gräns i rumslig upplösning som inte kan övervinnas. Flera super upplösning avbildningsmetoder har nyligen utvecklats för att kringgå denna gräns. En del bygger på lokalisering precision, till exempel stokastiska optisk rekonstruktion mikroskopi (STORM) <sup> 23,24, foto lokalisering mikroskopi (PALM) 25, fluorescens PALM (FPALM) 26, och single-partikel tracking PALM (sptPALM) 27: den relativt stora mängd fotoner som krävs på varje stillbild, men begränsar den tid upplösning dessa metoder till åtminstone flera millisekunder, vilket hämmar deras tillämplighet in vivo.
Däremot har ett lovande alternativ för superupplösning avbildning öppnats av rumsligt modulera fluorescens emission med stimulerade utarmning utsläppsmetoder (STED eller reversibla mätt optiska fluorescens övergångar (RESOLFT)) 28,29. Dessa tillvägagångssätt kombinerar utformningen av observationsvolymen långt under diffraktionsgränsen med möjlighet att använda snabb scanning mikroskop och detekteringssystem. I kombination med fluorescensfluktuationer analys, STED mikroskopi tillåtet att direkt söka av nanoskala Spatiotemporal dynamik lipider och proteins i levande cellmembran 30,31.
Samma fysiska kvantiteter av STED baserad mikroskopi kan erhållas genom en modifierad tid och rum bildkorrelationsspektroskopi (STICS 32,33) metod som är lämplig för att studera dynamiken i fluorescerande-märkta membranproteiner och / eller lipider i levande celler och genom en kommersiell mikroskop. Den experimentella protokollet presenteras här består av några få steg. Den första kräver en snabb avbildning av regionen av intresse på membranet. Därefter resulte trave bilder används för att beräkna den genomsnittliga rumsliga-temporala korrelationsfunktioner. Genom att montera den serie av korrelationsfunktioner, kan den molekylära "diffusion lag" erhållas direkt från avbildning i form av en skenbar diffusivitet (D app) – vs -Genomsnittlig förskjutning tomt. Denna tomt är kritiskt beroende miljön utforskas av molekylerna och tillåter att erkänna direkt själva spridningssättav lipid / protein av intresse.
I mer detaljer, som tidigare visats 34, den tid och rum autokorrelationsfunktion för den förvärvade bildserie beror kritiskt på dynamiken i molekylerna rör sig i den insamlade bildserien (observera att samma resonemang kan tillämpas på en rad förvärv där bara en dimension i rymden anses). Särskilt definierar vi korrelationsfunktionen som:
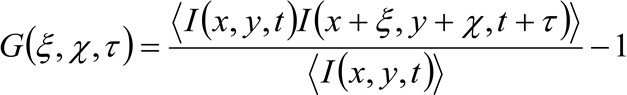 (1)
(1)
där 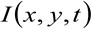 representerar den uppmätta fluorescensintensiteten i den position x, y och vid tiden t,
representerar den uppmätta fluorescensintensiteten i den position x, y och vid tiden t, ![]() och
och ![]() representerar avståndet i x-ochy-riktningarna respektive,
representerar avståndet i x-ochy-riktningarna respektive, ![]() representerar viss tid och
representerar viss tid och ![]() representerar genomsnittet. Denna funktion kan uttryckas som:
representerar genomsnittet. Denna funktion kan uttryckas som:
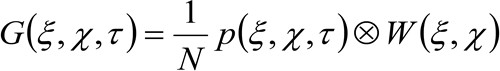 (2)
(2)
där "N" representerar det genomsnittliga antalet molekyler i området observation, ![]() representerar konvolution drift i rymden, och
representerar konvolution drift i rymden, och 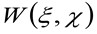 representerar autokorrelationen för den instrumen midjan. Detta senare kan tolkas som ett mått på hur fotoner från en enda sändare är utspridda i rymden på grund av att den optiska / inspelning setup (den så kallade punktspridningsfunktionen, PSF, genrally väl approximeras med en Gauss-funktion). Slutligen
representerar autokorrelationen för den instrumen midjan. Detta senare kan tolkas som ett mått på hur fotoner från en enda sändare är utspridda i rymden på grund av att den optiska / inspelning setup (den så kallade punktspridningsfunktionen, PSF, genrally väl approximeras med en Gauss-funktion). Slutligen 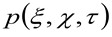 representerar sannolikheten för att finna en partikel på ett avstånd
representerar sannolikheten för att finna en partikel på ett avstånd ![]() och
och ![]() efter en tidsfördröjning
efter en tidsfördröjning ![]() . Om vi betraktar en diffusiva dynamik, där partiklar rör sig slumpmässigt i alla riktningar och nettoflöden inte är närvarande, är denna funktion också väl approximeras med en Gauss-funktion där variansen kan identifieras som Mean Square Displacement (MSD) i rörliga partikeln . Sålunda midjan på korrelationsfunktionen (även kallat
. Om vi betraktar en diffusiva dynamik, där partiklar rör sig slumpmässigt i alla riktningar och nettoflöden inte är närvarande, är denna funktion också väl approximeras med en Gauss-funktion där variansen kan identifieras som Mean Square Displacement (MSD) i rörliga partikeln . Sålunda midjan på korrelationsfunktionen (även kallat ![]() ), Kan definieras som summan av de partikel muskuloskeletala sjukdomar och den instrument midja och kan mätas genom en gaussisk passformting av korrelationsfunktionen för varje tidsfördröjning. Den uppmätta i MSD kan användas för att beräkna en skenbar diffusiviteten hos de rörliga molekyler
), Kan definieras som summan av de partikel muskuloskeletala sjukdomar och den instrument midja och kan mätas genom en gaussisk passformting av korrelationsfunktionen för varje tidsfördröjning. Den uppmätta i MSD kan användas för att beräkna en skenbar diffusiviteten hos de rörliga molekyler ![]() och en genomsnittlig förskjutning
och en genomsnittlig förskjutning ![]() som:
som:
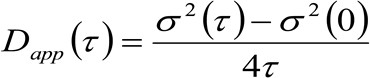 (3)
(3)
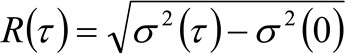 (4)
(4)
Några överväganden på experimentuppställning som används kan vägleda läsaren genom de följande avsnitten. För att selektivt excitera fluoroforema på basalmembranet av levande celler som vi kommer att använda en total inre reflektion (TIR) belysning, med hjälp av ett kommersiellt TIR fluorescens (TIRF) mikroskop (detaljer finns i materialet avsnitt). Dessutom, för att samla in the fluorescens vi kommer att använda en hög förstoring mål (100X NA 1,47, är hög numerisk apertur som krävs för TIRF belysning) och en EMCCD kamera (fysiska storleken på pixel på chipet 16 um). För att nå en pixelstorlek på 100 nm tillämpar vi en extra förstoringslins 1,6X. Såsom diskuteras nedan, skulle en tidsupplösning under en ms krävas för att korrekt beskriva dynamiken i snabba membranlipider nedanför 100 nm. För att uppnå denna tidsupplösning måste vi välja ett område av intresse (ROI) som är mindre än hela chipet av kameran (512 x 512). På detta sätt kommer kameran läsa ett reducerat antal linjer ökande tidsupplösningen. Men i denna avläsning regim ramtiden skulle begränsas av den tid som krävs för att flytta de avgifter från exponering för utläsningschipset på kameran och är oftast i storleksordningen millisekunder för 512 x 512 pixel EMCCD. För att slå denna gräns, kan en ny teknik flytta ROI-linjerna i stället för hela ramen, wed en praktisk effektiv minskning av den exponerade chip storlek (kallas Cropped Sensor läge i vårt EMCCD). För denna konfiguration för att vara effektiv, måste chipet utsidan av ROI täckas av ett par slitsar som är monterade i den optiska banan. Tack vare denna inställning en tidsupplösning ned till 10 -4 sekunder kan uppnås. Observera dock att detta tillvägagångssätt kan kopplas till många olika experimentella uppställningar, som förklaras i "diskussionen" avsnittet.
Demonstration av metoden kommer att tillhandahållas i levande celler, genom att använda både en ATTO488 märkt 1-palmitoyl-2-hydroxi-sn-glycero-3-fosfoetanolamin (ATTO488-PPE) och en GFP-märkt variant av transferrinreceptorn (GFP- TfR). I fallet med ATTO488-PPE kan detta tillvägagångssätt lyckades återställa en nästan konstant D appen som en funktion av genomsnittlig förskjutning indikerande en mestadels fri diffusion, såsom tidigare rapporterats 30,35. Däremot visar TfR-GFP en minskande D <sub> app som funktion av genomsnittlig förskjutning, vilket tyder delvis begränsad diffusion 6. Dessutom är det i det senare fallet är möjligt att kvantifiera den lokala diffusionskonstanten och genomsnitts inneslutning område under många mikrometer på membranplanet.
