To characterize histo-pathological features of SB-induced glioblastomas, C57/ BL6 neonatal mice were injected at P1 with a plasmid encoding luciferase (pT2/SB100x-Luc) in combination with plasmids encoding transposons with oncogenic DNA, i.e., NRAS (pT/CAGGS-NRASV12) and SV40 LgT (pT/CMVSV40-LgT) (Figure 3c) or a plasmid encoding a short hairpin p53 with PDGFβ and a GFP reporter (pT2shp53/GFP4/mPDGFβ) in combination with NRAS (Figure 3d). Animals were monitored for bioluminescence the day after injection (Figure 2a) and periodically until reaching the moribund stage when they were euthanized. Moribund stage is defined as the clinical stage when the animal becomes symptomatic showing impaired mobility, hunched posture, scruffy fur and weight loss. Sometimes, animals develop seizures or abnormal patterns of movement, like walking in circles and sudden jumping. At the end of the experiment animals were anesthetized. The brains were perfused, embedded in paraffin, and processed for hematoxylin and eosin staining. Data show tumors displaying the hallmarks of human GBM (WHO grade IV) with hemorrhages (Figure 3c), pseudo-pallisading necrosis (Figure 3e) and perivascular and diffuse invasion into the brain parenchyma (Figure 3g, f). The formation of pseudopallisades is preceded by the rupture of large glomeruloid vessels with leaky endothelia (Figure 3i) which result in regions of hemorrhage with massive infiltration of mononuclear cells(Figure 3h). Atypical mitoses (Figure 3j) and gigantic multinucleated tumor cells (Figure 3k) are also pathognomonic of human GBM.
Glioblastomas generated using the Sleeping Beauty transposase system can be monitored throughout the progression of tumor growth with bioluminescence, if the plasmids injected encode luciferase. An example experiment is shown in Figure 4a. Animals usually succumb of tumor burden when luminescence reaches an intensity of 107-109 photons/s/cm2/sr. The median survival of animals is predictably dependent on the combinations of oncogenic transposons injected into the neonatal brain, as illustrated in the survival curves presented in Figure 4b. Note that the most aggressive tumors are induced with NRAS and SV40 LgT antigen (median survival of 30 days), whereas the median survival of animals with GBM induced with shp53 NRAS and PDGF is 62.5 days and of animals injected with shp53 and NRAS 83 days.
De novo formed tumors can be characterized immuno-histochemically by their expression of molecules characteristic of glial tumors. Figure 5 shows a nascent tumor (22 dpi, bioluminescence 2×105 photons/s/cm2/sr), induced with shp53 and NRAS. The injected shp53 plasmid encodes for green fluorescent protein (GFP) to allow the identification of transfected cells and their progeny (pT2/shp53/GFP). GFP+ tumor cells express the neural stem cell marker nestin and some GFP+ cells also express glial fibrillary acidic protein (GFAP). Figure 6 illustrates a tumor from a moribund animal at 56 dpi, tumor which was also induced with shp53/GFP and NRAS. Numerous GFAP+ astrocytes are surrounding the tumor. GFP+ tumor cells express nestin, but not GFAP.
A great advantage when using this technique to induce GBMs is the ability to generate novel GBM cell lines with unique genetic alterations by means of the transposons injected. In addition, custom cell lines can be generated using transgenic animals with specific genetic makeup. These cell lines are instrumental in asking many mechanistic questions using biochemical assays. They are ideally suited for cytotoxicity studies with novel chemotherapeutic agents. Figure 7 illustrates a neurosphere from a Sleeping Beauty tumor induced with shp53, PDGF and NRAS, showing expression of GFP, which is encoded on one of the injected plasmids. Note that expression is not equally intense in all the cells, indicating the heterogeneous nature of these primary GBM cells.
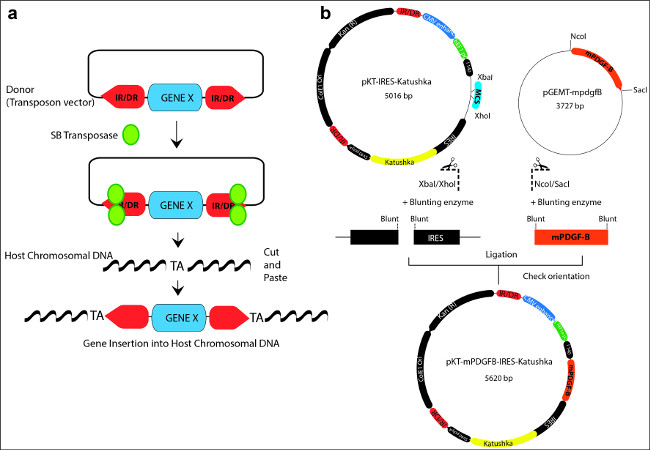
Figure 1: (a) Schematic representation illustrating the “cut and paste” mechanism used by the Sleeping Beauty transposase to integrate transposons into the host chromosomal DNA. A donor transposon plasmid is depicted with the gene of interest flanked by the inverted repeats/direct repeats (IR/DR; red arrow boxes) sequences. The SB transposase (green) binds to the IR/DR, excises the transposon and reintegrates it in between random TA dinucleotide base pairs on the host chromosomal DNA. (b) Example of cloning a gene of interest (mPDGFβ) into the backbone of a SB vector (pKT-IRES-Katushka). The SB vector contains two IR/DR repeat sequences (red) and a gene expression cassette which includes promoter and enhancer sequences, a multiple cloning site (MCS; blue), internal ribosomal entry sites (IRES), a fluorescent reporter marker (Katushka: yellow), and a polyadenilation site (polyA). The oncogene of interest (mPDGFβ; orange) is contained in a cloning vector pGEMT. The oncogene, flanked by specific restriction sites (NcoI/SacI) is released by enzymatic digestion, and blunted by the blunting enzyme. The vector is linearized and blunted as well. Finally, the mPDGFβ oncogene is inserted into the donor vector by means of a blunt-end ligation reaction to generate the new plasmid transposon vector: pKT-mPDGFβ-IRES-Katushka. Please click here to view a larger version of this figure.

Figure 2: Experimental Setup and guides for intra ventricular injections in neonatal mice (a) A stereotaxic frame (2) with micrometer dials is fitted with an automatic injector holding a 10 µl syringe (4). Inside the U frame, a neonatal adaptor frame is securely fastened (3). The control panel of the automatic injector (1) allows for precise selection of syringe, volume and rate of flow. (b) Photograph of a neonatal mouse (P1) with a needle inserted at the coordinates required for injections into the lateral ventricle: 1.5 mm ventral and 0.8 mm lateral to the lambda. (c) Illustration of a coronal section through the brain of a neonatal mouse (P1) highlighting the relative dimensions and position of the ventricles.

Figure 3: Tumors induced with the Sleeping Beauty transposon system show the histological hallmarks of human GBM (a) Bioluminescence image of a neonatal pup 24 hr after intraventricular injection with plasmids encoding NRAS, SV40 LgT. (b) Bioluminescence image of an adult animal (50 dpi) with a large tumor induced with shp53 NRAS and PDGF. (c) Hematoxilin and eosin stain of a coronal section from a brain of a moribund mouse with a tumor induced with NRAS and LgT (28 dpi). (d) Coronal section from a brain of a moribund mouse with a tumor induces with shp53 NRAS and PDGF. (e) Pseudopalisading necrosis, histological hallmark of human GBM is observed in de novo generated tumors. Arrow points to cells arranged in palisades migrating away from the central area of necrosis (N) (f) and (g) de novo generated tumors are highly invasive, showing invasion along blood vessels (g) and diffuse (f) into the normal brain parenchyma. (h) Region of hemorrhage with massive invasion of mononuclear cells (arrow), the initial stage of an area of pseudopalisading necrosis. (i) Large glomeruloid vessel with leaky endothelium (arrow) at the origin of diffusing hemorrhage inside a tumor induced with NRAS and SV40 –LgT. (j) Atypical mitosis in a tumor cells (arrowhead). (k) Gigantic tumor cell with multiple large nuclei (arrowhead).
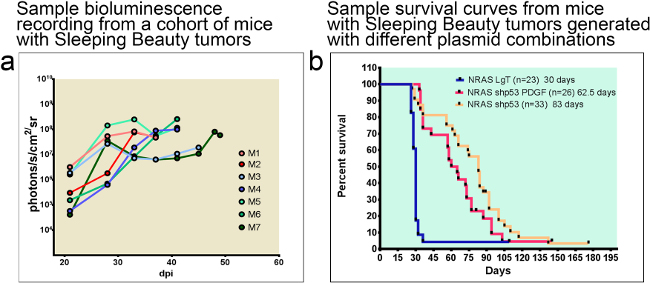
Figure 4: Tumor-bearing animals can be monitored with bioluminescence throughout the duration of disease progression. The median survival of animals is predicted by the combination of oncogenic DNA injected. (a) Example of bioluminescent traces from a cohort of 7 C57/BL6 mice. Note that mice succumb about a week after bioluminescence reaches 108 photons/s/cm2/sr. (b) Sample survival curves comparing median survival of animals with Sleeping Beauty tumors generated with different plasmid combinations. Median survival of tumors induced with NRAS and SV40 LgT is 30 dpi whereas of animals with tumors induced with shp53 NRAS and PDGF or shp53 and NRAS is 62.5 dpi or 83 dpi respectively. dpi: days post injection.
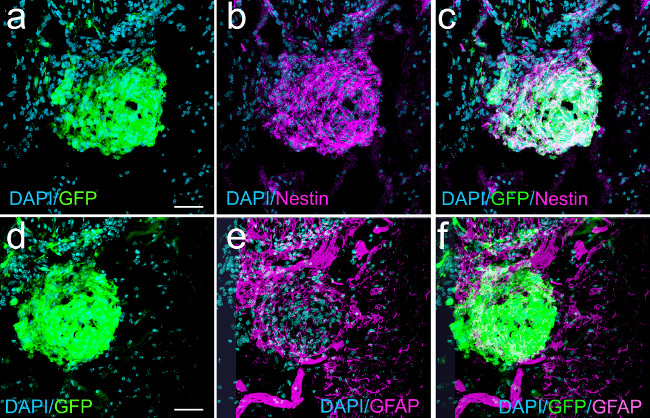
Figure 5: Nascent macroscopic GBM (22 dpi) induced with shp53 and NRAS show expression of nestin and GFAP in GFP+ tumor cells (confocal micrographs). Panel (a) shows a nascent tumor expressing GFP. Panel (b) represents the same field showing nestin expression. Panel (c) is an overlay of (a) and (b) illustrating co-localization of nestin and GFP. (d) GFP expression in tumor cells. (e) GFAP expression in some tumor cells and in cells surrounding the nascent tumor. (f) Overlay projection of (d) and (e) illustrating some tumor cells (white) co-expressing GFAP and GFP. Scale bars in (a) and (d) represent 75 µm.

Figure 6: GBM induced with shp53, NRAS and PDGF from a moribund animal (56 dpi) showing GFP and nestin expression in tumor cells and abundant staining for the mature glial marker GFAP surrounding the tumor. Panel (a) represents Nissl stain of acoronal section through the brain of a moribund animal with a tumor (intense blue staining from increased cellularity) induced with the Sleeping Beauty transposon system using shp53, PDGFΒ and NRAS. Panel (b), a coronal adjacent section to the one illustrated in panel (a) stained with the nuclear stain DAPI, showing expression of GFP in tumor cells. Panel (c) represents a confocal micrograph of the tumor border showing GFP expression in the tumor, identified by the high nuclear density with DAPI. Panel (d) illustrates the same field of view as in (a), showing intense GFAP expression in cells adjacent to the tumor. Panel (e) represents the overlay of panels (c) and (d). Panel (f) is a confocal micrograph of the tumor border, showing GFP expression in tumor cells. Panel (g) represents the same field of view as in (f), showing expression of nestin in tumor cells. Panel (h) is the overlay projection of panels (f) and (g). Scale bar in (c) represents 150 µm and in (f) represents 75 µm.

Figure 7: Neurospheres from a Sleeping Beauty tumor induced with shp53, PDGF and NRAS after 5 days in culture, passage 2. The cells are expressing GFP encoded on the shp53 PDGF plasmid (pT2shp53/GFP4/mPDGFβ). Panel (a) is a brightfield micrograph of a neurosphere. Panel (b) represents an epi-fluorescent micrograph of the same view as in (a) showing expression of GFP in the neurosphere cells. Panel (c) represents the overlay of panels (a) and (b).
