Membraan fusie is een universeel biologisch proces nodig is voor intracellulaire handel in vetten en eiwitten, secretie, de bevruchting, de ontwikkeling en omhuld virus binnenkomst in gastheerorganismen 1-3. Voor de meeste intracellulaire fusiereacties waaronder afgifte van hormonen en neurotransmitters via exocytose wordt de energie twee lipide bilagen smelten door vorming van een vier-helix bundel tussen cognate oplosbare N-ethylmaleimide-gevoelige factor attachment eiwitreceptor (SNARE) eiwitten, verankerd in het blaasje (v-SNARE) en het doelmembraan (t-SNARE) 4 resp. Synaptische vesikel exocytose is de meest strak gereguleerd fusiereactie en voordat één milliseconde na aankomst van een actiepotentiaal 1,4,5. De fusie porie, de initiële verbinding tussen de twee compartimenten fuseren, kunnen flikkeren open en meerdere malen gesloten voordat het opnieuw afsluiten of het uitbreiden van onomkeerbaar 5-7. De voormalige resultatenin voorbijgaande, "kiss & run" fusion, terwijl de laatste leidt tot volledige fusie. Factoren die het evenwicht tussen deze twee modi van fusie en mechanismen reguleren poriën flikkeren zijn niet goed begrepen 5,8.
SNARE eiwitten zijn nodig voor exocytose; synaptische blaasje fusie wordt na splitsing van SNAREs afgeschaft door neurotoxins 9. Bulk fusie-experimenten met kleine unilamellaire vesikels (SUV's) aangetoond dat SNAREs niet alleen vereist, maar ook voldoende membraanfusie 10 rijden. In deze massa assay SUV gereconstitueerd met v-SNAREs (v-SUV) werden gedoteerd met fluorescente fosfolipiden (N – (7-nitro-2-1,3-benzoxadiazol-4-yl) -phosphoethanolamine (NBD-PE) en ( N -. (lissamine rhodamine B sulfonyl) -phosphoethanolamine (LR-PE) en gemengd met ongelabeld blaasjes met t-SNAREs (t-SUV) Aanvankelijk was de fluorescentie van NBD-PE in v-SUV is geblust door Förster resonance energy transfer (FRET ) naar LR-PE. Zoals labeLED v-SUV fuseren met ongelabelde t-SUV, is de fluorofoor oppervlaktedichtheid in het nu gecombineerde membraan verminderd en de resulterende toename van NBD-PE fluorescentie meldt de mate van lipide mengen 10. Aangezien het grootste deel test is eenvoudig in te stellen en te analyseren, is het op grote schaal gebruikt om de mechanismen van SNARE gemedieerde fusie 10-14 bestuderen. Het heeft echter verscheidene beperkingen, zoals lage gevoeligheid en slechte tijdsresolutie. Belangrijker, als ensemble metingen is gemiddelde resultaten voor alle gebeurtenissen maken onderscheid tussen docking en fusion, alsmede opsporing van hemifusion tussenproducten moeilijk.
De afgelopen tien jaar verschillende groepen, waaronder de onze, zijn er nieuwe tests ontwikkeld om fusie gebeurtenissen bij de enkele blaasje level 15-27 volgen. Ha en collega's gebruikt v-SUV vastgebonden op een oppervlak en bewaakt hun fusie met gratis t-SUV 18,19. Lipide mengen werd gevolgd met behulp van FRET tussen twee lipide gebonden fluoroforen embedden in de v- en t-SUV's, respectievelijk, met behulp van totale interne reflectie fluorescentie (TIRF) microscopie 18. Later, Brunger het laboratorium gebruikt een enkele lipide-label soorten samen met een inhoud marker voor gelijktijdige detectie van lipiden en inhoud te mengen 20,28. Zowel de lipide en de inhoud merkers werden opgenomen in hoge zelfdovende concentraties; fusie met ongelabelde SUV resulteerde in fluorescentie dequenching 20,28.
Anderen hebben v-SUV gefuseerd met vlakke dubbellagen opgelost in t-SNAREs 15-17,21-27,29. De vlakke geometrie van het doel (t-SNARE bevattende) dubbellaag beter bootst de fysiologische fusieproces van kleine, zeer gebogen blaasjes met een platte plasmamembraan. De Steinem groep toegepast poriën spanning membranen opgelost in t-SNAREs, opgehangen boven een poreus siliciumnitride substraat en gedetecteerd fusion individuele v-SUV met behulp van confocale laser scanning microscopie 23. anderen fgebruikte v-SUV's naar vlakke dubbellagen opgelost in t-SNAREs, ondersteund op een glazen substraat 15-17,21,22,24-27,29. Het grote voordeel van het gebruik van ondersteunde bilagen (SBLs) dat TIRF microscopie kan worden gebruikt voor docking en fusiegebeurtenissen met uitstekende signaal-ruisverhouding en zonder interferentie van vrije v-SUV detecteren, hoewel behulp microfluïdische ook resolutie eenmalige gebeurtenis gebruikt standaard ver-field epifluorescentie microscopie 24.
Een belangrijk punt van zorg is de vraag of en hoe substraat-bilaag interacties invloed op ondersteunde bilaag kwaliteit en het fusieproces. Vroeg werk gebruik gemaakt van vlakke SBLs die direct werden ondersteund op een glazen of kwarts substraat 15-17. Deze SBLs werden door adsorptie, barsten, verspreiding en fusie van t-SUV membranen op het substraat. Het werd al snel besefte echter dat het weglaten van een belangrijke t-SNARE component, SNAP25, van SBLs op deze manier bereid resulteerde in v-SUV docking en fusion kinetiek indistinguishable van die verkregen met een totale t-SNAREs 17. Omdat SNAP25 absoluut voor fusie is nodig in vivo 30,31, werd de fysiologische relevantie van deze vroege pogingen in twijfel te trekken. Tamm de groep overwon deze uitdaging door het gebruik van beter gecontroleerd ondersteunde bilaag vorming 21. Vroeger Langmuir-Blodgett depositie voor het eiwit-vrij eerste folder van de SBL, gevolgd door de fusie van die monolaag met t-SUV's 21. Dit resulteerde in SNAP25-afhankelijke fusie.
Om potentiële artefacten geassocieerd met een dubbellaag direct afgezet op een glassubstraat zonder nader Langmuir-Blodgett methoden voorkomen Karatekin et al. Werd een zachte, gehydrateerde poly (ethyleenglycol) (PEG) kussen tussen de bilaag en het substraat 24. Deze modificatie resulteerde ook in SNAP25-afhankelijke fusie 24. Dubbellagen kussens op een zachte polymeerlaag was gekend om transmembraan beter behoudenmobiliteit en de functie 32-eiwit, en was gebruikt in fusion studies met virussen 33. Bovendien gePEGyleerd dubbellagen lijken wat vermogen om zichzelf te genezen behouden en zijn zeer robuust 34,35. Eerst wordt een fractie van commercieel verkrijgbare, lipide-gebonden PEG-ketens zijn opgenomen in de t-SUV membraan. Wanneer deze t-SUV burst en vormen een vlakke dubbellaag op een glazen substraat, een PEG borstel omvat zowel bladen van de vlakke bilaag. Omdat vorming vlakke dubbellaag wordt aangedreven door adhesie van het PEG-ketens rond t-SUV op het hydrofiele glasoppervlak, liposoom barsten en vlakke bilaag vorming betrekkelijk ongevoelig voor de lipidesamenstelling gebruikt. Wanneer echter grote hoeveelheden cholesterol zijn opgenomen, waardoor de cohesieve eigenschappen van de SUV, de SUV kan niet spontaan barsten. Als dit het geval is, kan osmotische shock of tweewaardige ionen worden toegepast om vlakke bilaag vorming 25 helpen.
Zoals hierboven vermeld, in dit apdering een PEG borstel omvat beide zijden van de vlakke, ondersteund dubbellaag. De borstel tegenover de microfluïdische stromingskanaal helpt om niet-specifieke hechting van inkomende v-SUV die ook meestal bedekt met een PEG-laag voorkomen. Vorming van V- en t-SNARE complexen begint vanaf de membraan-distale N-uiteinden en verloopt in etappes in de richting van het membraan-proximale domeinen 36. Voor de v-SUV om met de t-SBL, V- en t-SNARE N-termini nodig boven het PEG borstels, die lijkt op het geval onder de omstandigheden van de assay uitsteken. Borstelhoogte kan worden aangepast aan andere eiwitten dan SNAREs bestuderen door het variëren van de dichtheid van gepegyleerde lipiden en PEG ketenlengte 37,38. Een ander voordeel van de PEG borstels die de proximale oppervlakken van het fuseren van bilagen is dat zij bootsen de drukke omgeving van biologische membranen die zijn verpakt met 30.000-40.000 integrale membraaneiwitten per vierkante micron 39. Net als de PEG-ketens in deze test, de Repulsive eiwit laag die biologische membranen moeten opzij worden geduwd om te zorgen voor contact tussen de twee fosfolipiden dubbellagen voor fusie te komen.
Microfluïdische stromingskanalen worden in deze test, omdat ze unieke voordelen bieden. Ten eerste microfluïdische stroom kunnen meer uniforme afzetting van t-SUV te verspreiden en de zekering t-SBL vormen. Ten tweede, de kleine kanaal volume (<1 pl) minimaliseert monster consumptie. Ten derde, de kleine volumina vereist waarvan de hele experiment constant debiet te worden uitgevoerd. Stroom verwijdert zwak, vermoedelijk aspecifiek, gehecht v-SUV van SBL 16. Het heeft ook een constante dichtheid van v-SUV boven de t-SBL, het vereenvoudigen van kinetische analyse 17. Tenslotte worden gedokt blaasjes gemakkelijk te onderscheiden van vrije degenen door de stroming 25 uitgevoerd. Ten vierde, meerdere microkanalen kunnen worden gebruikt op dezelfde dekglaasje, elk probing een andere toestand. Dit maakt vergelijking van de voorwaarden During dezelfde experimentele run. Een soortgelijke aanpak is gebruikt door de van Oijen groep fusie tussen influenzavirus en kussens SBLs 33 bestuderen.
Bij TIRF microscopie, de exponentiële verval van het verdwijnende veld (met vervalconstante ~ 100 nm) fluorescentie excitatie beperkt tot die moleculen die in de directe nabijheid van het glas-buffer interface. Dit minimaliseert bijdrage van fluorescente moleculen die verder weg zijn, verhoogt de signaal-ruisverhouding en maakt enkel molecuul gevoeligheid belichtingsraam tijden van 10-40 msec. Het verdwijnende veld leidt ook tot een toename van signaal-fusie: de overdracht gemerkte lipiden uit de SUV in de SBL, bevinden zij zich gemiddeld sterker bekrachtigingsveld. Deze toename in fluorescentie wordt sterker grotere liposomen.
Als gepolariseerd licht wordt gebruikt om het verdwijnende veld genereren extra effecten bijdragen tot een verandering in fluorescentie bij transfer van etiketten van de SUV in de SBL. Sommige lipide kleurstoffen overgang dipool georiënteerd met een voorkeurs gemiddelde hoek ten opzichte van de dubbellaag waarin zij zijn ingebed. Hierdoor ontstaat een verschil in de hoeveelheid fluorescentie die door de fluoroforen wanneer zij in de SUV versus de SBL, aangezien het gepolariseerde bundel kleurstoffen anders prikkelen de twee membranen. In het eerste geval zal de excitatie balk interactie met overgang dipolen georiënteerde rond de sferische SUV, terwijl voor het laatste, zal dipool oriëntaties worden beperkt door de vlakke SBL geometrie. Wanneer bijvoorbeeld s-gepolariseerd invallend licht (gepolariseerd loodrecht op het vlak van inval) wordt gebruikt, excitatie efficiënter als de kleurstof in de SBL dan in SUV voor een lipide kleurstof transitiedipool evenwijdig aan het membraan 29,40 (zoals die van Dil of heb 41-43). Een SUV gedoteerd met zo'n fluorofoor verschijnt dimmen wanneer het dokken op de SBL (figuur 7, Representat ive resultaten). Als een fusie porie opent en verbindt de SUV en SBL membranen, fluorescerende probes diffunderen in de SBL en meer kans te worden enthousiast over de-s gepolariseerde verdwijnende veld 25,27,29. Bijgevolg het fluorescentiesignaal geïntegreerd rondom de fusieplaats sterk toeneemt tijdens de kleurstofoverdracht van de SUV in de SBL 27 (Figuur 3 en Figuur 7). Een bijkomende factor die bijdraagt aan veranderingen die gepaard fusie wordt dequenching van fluorescente labels zoals ze worden verdund wanneer overgebracht in de SBL signaal. De bijdrage van dequenching gewoonlijk klein in vergelijking met verdwijnende veld verval en polarisatie-effecten in de hier beschreven test, omdat slechts een kleine fractie ( 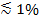 ) Van de lipiden zijn gelabeld.
) Van de lipiden zijn gelabeld.
Het signaal stijging op fusie kan worden benut om fusie porie eigenschappen afleiden door de tijd te vergelijken,1 "src =" / files / ftp_upload / 54349 / 54349eq2.jpg "/>, die nodig is voor een lipide te ontsnappen via een porie die vrij doorlaatbaar voor lipiden aan de werkelijke release tijd,  . Als de twee tijdschalen vergelijkbaar zou worden geconcludeerd dat de poriën weer weinig weerstand tegen lipide flow. Indien de werkelijke lostijd aanzienlijk langer is dan de tijd voor diffusie beperkte release, zou een proces aangeven, zoals poriën flikkeren, vertragen lipide release. De diffusie beperkte release tijd,
. Als de twee tijdschalen vergelijkbaar zou worden geconcludeerd dat de poriën weer weinig weerstand tegen lipide flow. Indien de werkelijke lostijd aanzienlijk langer is dan de tijd voor diffusie beperkte release, zou een proces aangeven, zoals poriën flikkeren, vertragen lipide release. De diffusie beperkte release tijd, 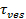 , Afhankelijk van de grootte van de fuser en liposoom lipide-diffusie; haar ramingen vereist deze twee parameters te worden gekwantificeerd. De single molecule gevoeligheid van de test maakt het mogelijk lipide diffusie te meten door het bijhouden van meerdere afzonderlijke lipide fluoroforen na hun vrijlating in het SBL voor elke fusie event 26. De grootte van elke versmelten vesiclekan worden geschat 27 door het combineren van (i) de intensiteit van een enkele lipide kleurstof, (ii) de verandering in de totale fluorescentie rond docking site na alle fluoroforen op fusie worden overgebracht in de SBL, (iii) de bekende etikettering dichtheid van SUV lipiden, en (iv) de oppervlakte per lipide. Voor veel fusiegebeurtenissen werden de werkelijke lipide afgifte samengesteld worden veel langzamer dan verwacht door diffusie gecontroleerde afgifte 27 zijn, zoals eerder werd opgemerkt uitgaande uniforme SUV maat 44. Uitgaande vertraging van lipide afgifte wordt veroorzaakt door het trillen poriën, een kwantitatief model maakt schatting van "porie openheid", de fractie van tijd de poriën blijft ook voor fusie-27.
, Afhankelijk van de grootte van de fuser en liposoom lipide-diffusie; haar ramingen vereist deze twee parameters te worden gekwantificeerd. De single molecule gevoeligheid van de test maakt het mogelijk lipide diffusie te meten door het bijhouden van meerdere afzonderlijke lipide fluoroforen na hun vrijlating in het SBL voor elke fusie event 26. De grootte van elke versmelten vesiclekan worden geschat 27 door het combineren van (i) de intensiteit van een enkele lipide kleurstof, (ii) de verandering in de totale fluorescentie rond docking site na alle fluoroforen op fusie worden overgebracht in de SBL, (iii) de bekende etikettering dichtheid van SUV lipiden, en (iv) de oppervlakte per lipide. Voor veel fusiegebeurtenissen werden de werkelijke lipide afgifte samengesteld worden veel langzamer dan verwacht door diffusie gecontroleerde afgifte 27 zijn, zoals eerder werd opgemerkt uitgaande uniforme SUV maat 44. Uitgaande vertraging van lipide afgifte wordt veroorzaakt door het trillen poriën, een kwantitatief model maakt schatting van "porie openheid", de fractie van tijd de poriën blijft ook voor fusie-27.
Wanneer praktisch is belangrijk om fusie mechanismen waarbij zowel lipide en oplosbare inhoud labels testen. Bijvoorbeeld zou lipide afgifte vertraagd worden door andere dan de porie trillen werkwijzen, zoals restrictie lipide diffusie door de SNARE eiwitten die surround thij porie. Als dit het geval zou zijn, dan los van de inhoud zou release van lipide labels voorafgaan, op voorwaarde dat de porie is groot genoeg om doorgang van oplosbare probes mogelijk te maken. Een fundamentele fout in de benadering kan in de veronderstelling dat de overdracht van gemerkte lipiden SBL vindt plaats via een smalle fusie porie verbinden de SBL een vesicle die grotendeels zijn pre-fusie-vorm heeft behouden. Lipid overdracht in de SBL kan ook het gevolg zijn van de snelle uitzetting van de fusie porie met een gelijktijdige, extreem snelle ineenstorting van de SUV in de SBL membraan, zoals eerder gesuggereerd op basis van lipide vrijlating data alleen 29. Monitoring zowel lipide en inhoud kort tegelijk bleek dat veel poriën afgedicht na het loslaten alle lipide labels, maar bleven sommige van hun oplosbare lading 27. Dit geeft aan dat ten minste enkele liposomen niet samen in de SBL na fusie en dat het lipide kleurstofoverdracht in de SBL gebeurt door een fusie porie. Bovendien, lIPID en inhoud vrijlating vond plaats gelijktijdig 27, waardoor het onwaarschijnlijk is dat de vertraging van lipide vrijlating was te wijten aan hinder van lipide diffusie door de SNARE eiwitten rond de porie 45.
Een SUV-SBL fusie-protocol dat niet controleren oplosbaar inhoud vrijlating is eerder gepubliceerd door Karatekin en Rothman 25. Hier worden meer recente ontwikkelingen opgenomen, namelijk de gelijktijdige controle van lipiden en de inhoud vrij te geven en de schatting van SUV, lipide, en fusie porie eigenschappen 27. Het protocol begint met instructies voor het bereiden van de microfluïdische cellen, gemaakt door het binden van een poly (dimethylsiloxaan) (PDMS) elastomeer blok met groeven met een glas dekglaasje 25. Vervolgens wordt de bereiding van v-SUV met zowel lipide en inhoud markers toegelicht. Afdelingen 4 en 5 geven instructies voor de montage van de microfluïdische cellen, de vorming van de SBLs in situ en het controleren op gebreken en vloeibaarheid, de invoering van vSUV's in de stroom cellen en detectie van fusiegebeurtenissen. Hoofdstuk 6 bevat instructies voor data-analyse.
