झिल्ली संलयन एक सार्वभौमिक जैविक प्रक्रिया लिपिड और प्रोटीन, स्राव, निषेचन, विकास के intracellular तस्करी के लिए आवश्यक है, और मेजबान जीवों 1-3 में वायरस प्रवेश छा। Exocytosis के माध्यम से हार्मोन और न्यूरोट्रांसमीटर की रिहाई सहित ज्यादातर intracellular संलयन प्रतिक्रियाओं के लिए, दो लिपिड bilayers फ्यूज करने के लिए ऊर्जा आत्मीय घुलनशील एन ethylmaleimide के प्रति संवेदनशील कारक लगाव प्रोटीन रिसेप्टर (जाल) प्रोटीन के बीच एक चार हेलिक्स बंडल के गठन के द्वारा प्रदान की जाती है, में लंगर डाले पुटिका (वि जाल) और लक्ष्य झिल्ली (टी-जाल) 4, क्रमशः। Synaptic पुटिका exocytosis सबसे कसकर विनियमित संलयन प्रतिक्रिया है और एक संभावित कार्रवाई 1,4,5 के आने के बाद एक millisecond के भीतर होता है। फ्यूजन ताकना, दो fusing डिब्बों के बीच प्रारंभिक कनेक्शन, खुले झिलमिलाहट और resealing या अचल 5-7 विस्तार करने से पहले कई बार बंद कर सकते हैं। पूर्व परिणामक्षणिक में, जबकि दूसरा सुराग पूर्ण विलय के लिए "चुंबन और भागो" संलयन। फ्यूजन के इन दो मोड और ताकना चंचल को विनियमित करने के तंत्र के बीच संतुलन शासी कारक अच्छी तरह से 5,8 समझ नहीं रहे हैं।
SNARE प्रोटीन exocytosis के लिए आवश्यक हैं; synaptic पुटिका संलयन न्यूरोटोक्सिन 9 से फन्दे की दरार पर समाप्त कर दिया है। थोक संलयन छोटे unilamellar पुटिकाओं (एसयूवी) का उपयोग कर प्रयोगों झिल्ली संलयन 10 ड्राइव करने के लिए पर्याप्त पता चला कि फन्दे केवल आवश्यक नहीं कर रहे हैं, लेकिन यह भी। इस थोक परख में, एसयूवी वी के फन्दे (वि एसयूवी) के साथ पुनर्गठन फ्लोरोसेंट फॉस्फोलिपिड (एन साथ डाल दिया गया गया है – (7-नाइट्रो-2-1,3-benzoxadiazol-4-YL) -phosphoethanolamine (NBD पीई) और ( एन -। (lissamine rhodamine बी Sulfonyl) -phosphoethanolamine (एलआर पीई) और युक्त टी फन्दे (टी एसयूवी) वी एसयूवी में NBD पीई के प्रारंभ में प्रतिदीप्ति Förster प्रतिध्वनि ऊर्जा हस्तांतरण से बुझती है लेबल हटाया गया पुटिकाओं के साथ मिश्रित (झल्लाहट ) एलआर-पीई के लिए। प्रयोगशाला के रूप मेंeLED वी एसयूवी लेबल हटाया गया टी एसयूवी के साथ फ्यूज, अब संयुक्त झिल्ली में fluorophore सतह घनत्व कम हो जाता है और NBD पीई प्रतिदीप्ति में जिसके परिणामस्वरूप वृद्धि लिपिड मिश्रण 10 की हद तक रिपोर्ट। थोक परख की स्थापना की और विश्लेषण करने के लिए आसान है, यह व्यापक रूप से जाल की मध्यस्थता संलयन 10-14 के तंत्र का अध्ययन करने के लिए इस्तेमाल किया गया है। हालांकि, यह इस तरह के कम संवेदनशीलता और गरीब समय संकल्प के रूप में कई सीमाओं की है। सबसे महत्वपूर्ण बात, एक जोड़ा माप के रूप में, मुश्किल hemifusion मध्यवर्ती की डॉकिंग और फ्यूजन के बीच भेदभाव, साथ ही पता लगाने कर रही सभी घटनाओं पर यह औसत का परिणाम है।
हमारा सहित पिछले एक दशक के कई समूहों, ओवर, एकल पुटिका स्तर 15-27 में संलयन घटनाओं पर नजर रखने के लिए नए assays विकसित किया है। हा और उनके सहयोगियों वी एसयूवी एक सतह पर सीमित इस्तेमाल किया और साथ मुफ्त T-एसयूवी 18,19 उनके संलयन पर नजर रखी। लिपिड मिश्रण लिपिड को बाध्य fluorophores की एक जोड़ी के बीच झल्लाहट का उपयोग करते हुए नजर रखी थी उन्हेंवि और टी-एसयूवी, क्रमशः में पलंगों वाले, कुल आंतरिक प्रतिबिंब प्रतिदीप्ति (TIRF) माइक्रोस्कोपी 18 का उपयोग कर। बाद में, Brunger की प्रयोगशाला के लिए एक एकल लिपिड लेबल प्रजातियां एक साथ इस्तेमाल किया लिपिड और सामग्री मिश्रण 20,28 के एक साथ पता लगाने के लिए एक सामग्री मार्कर के साथ। दोनों लिपिड और सामग्री मार्कर के उच्च, स्व-शमन सांद्रता में शामिल थे; लेबल हटाया गया एसयूवी के साथ फ्यूजन प्रतिदीप्ति में हुई 20,28 dequenching।
दूसरों टी फन्दे 15-17,21-27,29 साथ पुनर्गठन तलीय bilayers को वी-एसयूवी जुड़े हुए हैं। लक्ष्य (टी-जाल से युक्त) bilayer बेहतर mimics छोटे, एक फ्लैट प्लाज्मा झिल्ली के साथ अत्यधिक घुमावदार पुटिकाओं के शारीरिक संलयन की प्रक्रिया के तलीय ज्यामिति। Steinem समूह T-फन्दे साथ पुनर्गठन ताकना फैले झिल्ली, एक झरझरा सिलिकॉन नाइट्राइड सब्सट्रेट पर निलंबित कार्यरत हैं और लेजर confocal माइक्रोस्कोपी स्कैनिंग 23 का उपयोग कर व्यक्ति वी एसयूवी के साथ फ्यूजन का पता चला। दूसरों चटी फन्दे साथ पुनर्गठन तलीय bilayers करने के लिए खेतों में वी-एसयूवी, एक गिलास सब्सट्रेट 15-17,21,22,24-27,29 पर समर्थन किया। समर्थित bilayers (SBLs) का उपयोग कर के महान लाभ यह है कि TIRF माइक्रोस्कोपी, उत्कृष्ट संकेत करने वाली शोर अनुपात के साथ और से मुक्त V-एसयूवी हस्तक्षेप के बिना डॉकिंग और फ्यूजन की घटनाओं का पता लगाने के लिए इस्तेमाल किया जा सकता है, हालांकि microfluidics का उपयोग कर भी उपयोग कर एकल घटना संकल्प प्रदान करता है मानक दूर क्षेत्र epifluorescence माइक्रोस्कोपी 24।
एक प्रमुख चिंता का विषय है कि क्या और कैसे सब्सट्रेट-bilayer बातचीत bilayer गुणवत्ता और संलयन की प्रक्रिया का समर्थन किया प्रभावित करती है। प्रारंभिक काम है कि सीधे एक गिलास या क्वार्ट्ज सब्सट्रेट 15-17 पर समर्थित थे तलीय SBLs का इस्तेमाल किया। ये SBLs सोखना द्वारा किए गए थे, फोड़, प्रसार और सब्सट्रेट पर टी एसयूवी झिल्ली के संलयन। शीघ्र ही यह एहसास हो गया था कि, हालांकि, एक प्रमुख टी SNARE घटक को छोड़ते हुए, SNAP25, इस तरीके से तैयार SBLs से वी-एसयूवी डॉकिंग और फ्यूजन कैनेटीक्स indistinguis में हुईपूरा टी फन्दे 17 का उपयोग कर प्राप्त उन लोगों से Hable। क्योंकि SNAP25 बिल्कुल विवो 30,31 में संलयन के लिए आवश्यक है, इन शुरुआती प्रयासों की शारीरिक प्रासंगिकता सवाल में डाल दिया था। Tamm के समूह बेहतर समर्थित नियंत्रित bilayer गठन 21 का उपयोग करके इस चुनौती overcame। यह प्रोटीन से मुक्त एसबीएल के पहले पत्रक, टी 21 के साथ एसयूवी कि monolayer के विलय के बाद के लिए Langmuir-Blodgett बयान का इस्तेमाल किया। इस SNAP25 निर्भर संलयन में हुई।
एक bilayer सीधे Langmuir-Blodgett तरीकों का उपयोग करने की आवश्यकता के बिना एक गिलास सब्सट्रेट पर समर्थित के साथ जुड़े संभावित कलाकृतियों से बचने के लिए, Karatekin एट अल। शुरू की एक नरम, हाइड्रेटेड पाली (एथिलीन ग्लाइकोल) (खूंटी) bilayer और सब्सट्रेट 24 के बीच तकिया। इस संशोधन भी SNAP25 निर्भर फ्यूजन 24 में हुई। एक नरम बहुलक परत पर तकिये bilayers बेहतर transmembrane संरक्षित करने के लिए ज्ञात किया गया थाप्रोटीन गतिशीलता और समारोह 32, और वायरस के 33 के साथ फ्यूजन अध्ययन में इस्तेमाल किया गया था। इसके अलावा, PEGylated bilayers-चंगा आत्म करने के लिए कुछ करने की क्षमता बनाए रखने के लिए लग रहे हैं और बहुत मजबूत 34,35 हैं। सबसे पहले, व्यावसायिक रूप से उपलब्ध, लिपिड से जुड़े खूंटी चेन का एक अंश टी एसयूवी झिल्ली में शामिल किए गए हैं। इन टी-एसयूवी फट और एक गिलास सब्सट्रेट पर एक planar bilayer फार्म जब, एक खूंटी ब्रश तलीय bilayer के दोनों पत्रक शामिल किया गया है। क्योंकि तलीय bilayer गठन हाइड्रोफिलिक कांच की सतह पर आसपास के टी-एसयूवी खूंटी चेन के आसंजन द्वारा संचालित है, liposome फोड़ और तलीय bilayer गठन अपेक्षाकृत लिपिड इस्तेमाल किया संरचना करने के लिए असंवेदनशील हैं। हालांकि, जब कोलेस्ट्रॉल की बड़ी मात्रा में शामिल किए गए हैं, एसयूवी के एकजुट गुण बढ़ रही है, एसयूवी अनायास नहीं फट सकता है। यदि यह मामला है, आसमाटिक सदमे या द्विसंयोजक आयनों तलीय bilayer गठन 25 मदद करने के लिए नियोजित किया जा सकता है।
ऊपर उल्लेख किया है, इस एपी के रूप मेंproach एक खूंटी ब्रश तलीय के दोनों पक्षों को शामिल किया गया है, bilayer का समर्थन किया। ब्रश microfluidic प्रवाह चैनल का सामना करना पड़ भेजे वी एसयूवी जो भी आम तौर पर एक खूंटी परत के साथ कवर कर रहे हैं की अविशिष्ट आसंजन को रोकने में मदद करता है। वि और टी-जाल परिसरों के गठन झिल्ली बाहर एन Termini और झिल्ली समीपस्थ डोमेन 36 की ओर चरणों में आय से शुरू होता है। वी-एसयूवी टी एसबीएल, वि और खूंटी ब्रश, जो परख की शर्तों के तहत मामला प्रतीत हो रहा है ऊपर फैलाना टी SNARE एन Termini जरूरत के साथ बातचीत करने के लिए। ब्रश ऊंचाई PEGylated लिपिड का घनत्व और खूंटी श्रृंखला लंबाई 37,38 अलग से फन्दे से अन्य प्रोटीन का अध्ययन करने के लिए अनुकूलित किया जा सकता है। Fusing bilayers के समीपस्थ सतहों को कवर खूंटी ब्रश का एक अन्य लाभ यह है कि वे जैविक झिल्लियों जो वर्ग माइक्रोन 39 प्रति 30,000-40,000 अभिन्न झिल्ली प्रोटीन के साथ पैक कर रहे हैं के भीड़ भरे वातावरण की नकल है। बस इस परख में खूंटी चेन की तरह, repulsive प्रोटीन की परत जैविक झिल्लियों को कवर करने के लिए एक तरफ संलयन होने के लिए दो फॉस्फोलिपिड bilayers के बीच संपर्क के लिए अनुमति देने के लिए धक्का दिया जा जरूरत है।
Microfluidic प्रवाह चैनल, इस परख में इस्तेमाल के रूप में वे अद्वितीय लाभ की पेशकश कर रहे हैं। सबसे पहले, microfluidic प्रवाह टी एसबीएल फार्म के लिए टी-एसयूवी की अधिक समान बयान प्रसार करने के लिए और फ्यूज सक्षम बनाता है। दूसरा, छोटे चैनल मात्रा (<1 μl) नमूना खपत को कम करता है। तीसरा, छोटे आवश्यक मात्रा पूरे प्रयोग के निरंतर प्रवाह के तहत आयोजित किया जा करने के लिए अनुमति देते हैं। प्रवाह कमजोर निकालता है, शायद गैर-विशेष रूप से, एसबीएल 16 से पालन वी एसयूवी। यह भी टी एसबीएल ऊपर वी एसयूवी, गतिज विश्लेषण 17 को सरल बनाने की एक निरंतर घनत्व को बनाए रखता है। अंत में, डॉक पुटिकाओं आसानी से लोगों को मुफ्त प्रवाह 25 के द्वारा किया जाता से प्रतिष्ठित हैं। चौथा, कई microfluidic चैनलों ही coverslip पर इस्तेमाल किया जा सकता है, प्रत्येक एक अलग हालत की जांच कर रही। यह शर्तों Duri की तुलना की अनुमति देता हैएक ही प्रयोगात्मक रन एनजी। एक समान दृष्टिकोण वैन Oijen समूह द्वारा इस्तेमाल किया गया है इन्फ्लूएंजा वायरस और तकिये SBLs 33 के बीच विलय का अध्ययन करने के लिए।
TIRF माइक्रोस्कोपी में, क्षणभंगुर क्षेत्र का तेजी से क्षय उन अणुओं है कि गिलास बफर इंटरफेस के बहुत करीब निकटता में हैं सीमीत प्रतिदीप्ति उत्तेजना (एक क्षय निरंतर ~ 100 एनएम के साथ)। इस फ्लोरोसेंट अणुओं है कि आगे दूर कर रहे हैं के योगदान को कम करता है, संकेत करने वाली शोर अनुपात बढ़ जाती है, और 10-40 मिसे के फ्रेम जोखिम समय के साथ एक अणु संवेदनशीलता की अनुमति देता है। क्षणभंगुर क्षेत्र भी संलयन पर एक संकेत वृद्धि हो जाती है: एसबीएल में एसयूवी से लेबल लिपिड हस्तांतरण के रूप में, वे खुद को पाते हैं, औसतन, एक मजबूत उत्तेजना क्षेत्र में। प्रतिदीप्ति में यह वृद्धि बड़ा liposomes के लिए मजबूत है।
ध्रुवीकरण प्रकाश क्षणभंगुर क्षेत्र उत्पन्न करने के लिए प्रयोग किया जाता है, तो अतिरिक्त प्रभाव टी पर प्रतिदीप्ति में परिवर्तन करने के लिए योगदानएसबीएल में एसयूवी से लेबल की ransfer। कुछ लिपिड रंगों एक संक्रमण द्विध्रुवीय bilayer जिसमें वे एम्बेडेड रहे हैं करने के लिए सम्मान के साथ एक पसंदीदा मतलब कोण के साथ उन्मुख है। के बाद से ध्रुवीकृत किरण अलग ढंग से दो झिल्लियों में रंगों उत्तेजित होगा यह fluorophores द्वारा उत्सर्जित जब वे एसबीएल बनाम एसयूवी में हैं प्रतिदीप्ति की राशि में एक अंतर पैदा करता है। पूर्व के लिए, उत्तेजना बीम, गोलाकार एसयूवी चारों ओर उन्मुख संक्रमण द्विध्रुव के साथ बातचीत जबकि बाद के लिए, द्विध्रुवीय झुकाव द्वारा फ्लैट एसबीएल ज्यामिति ही सीमित हो जाएगा। उदाहरण के लिए, जब एस ध्रुवीकृत घटना प्रकाश (घटना के विमान को सामान्य ध्रुवीकरण) का इस्तेमाल किया जाता है, उत्तेजना और अधिक कुशल है जब डाई झिल्ली 29,40 करने के लिए एक लिपिड डाई संक्रमण द्विध्रुवीय उन्मुख समानांतर के लिए एसयूवी की तुलना में एसबीएल में है (जैसे DiI की है कि या था 41-43 के रूप में)। एक एसयूवी में इस तरह के एक fluorophore साथ डाल दिया गया मंद प्रकट होता है जब यह एसबीएल (चित्रा 7, REPRESENTAT पर नाव ive परिणाम)। एक संलयन रोमकूप खुल जाता है और एसयूवी और एसबीएल झिल्ली को जोड़ता है के रूप में, फ्लोरोसेंट जांच एसबीएल में फैलाना और अधिक एस ध्रुवीकरण क्षणभंगुर क्षेत्र 25,27,29 से उत्साहित होने की संभावना बन जाते हैं। नतीजतन, प्रतिदीप्ति संलयन साइट के आसपास एकीकृत संकेत एसबीएल 27 (3 चित्रा और चित्रा 7) में एसयूवी से डाई हस्तांतरण के दौरान तेजी से बढ़ जाती है। एक अतिरिक्त कारक है कि परिवर्तन के साथ फ्यूजन फ्लोरोसेंट लेबल के dequenching है के रूप में वे पतला कर रहे हैं जब एसबीएल में स्थानांतरित संकेत करने के लिए योगदान देता है। dequenching के योगदान, परख यहाँ वर्णित में क्षणभंगुर क्षेत्र क्षय और ध्रुवीकरण प्रभाव की तुलना में आम तौर पर नाबालिग है, क्योंकि केवल एक छोटा सा अंश ( 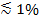 ) लिपिड के लेबल रहे हैं।
) लिपिड के लेबल रहे हैं।
फ्यूजन पर संकेत वृद्धि समय की तुलना द्वारा संलयन ताकना गुण परिणाम निकालना शोषण किया जा सकता है,1 "src =" / files / ftp_upload / 54349 / 54349eq2.jpg "/>, एक लिपिड एक ताकना कि स्वतंत्र रूप से वास्तविक रिलीज के समय के लिए लिपिड के लिए पारगम्य है के माध्यम से बचने के लिए आवश्यक है,  । दो समय तराजू तुलना कर रहे हैं, तो यह निष्कर्ष निकाला जा सकता है कि ताकना लिपिड प्रवाह करने के लिए थोड़ा प्रतिरोध प्रस्तुत करता है। हालांकि, अगर वास्तविक रिलीज के समय में काफी प्रसार सीमित रिहाई के लिए समय की तुलना में अब है, यह एक प्रक्रिया ऐसी ताकना चंचल के रूप में संकेत होता है, लिपिड रिहाई की गति को धीमा। प्रसार-सीमित रिलीज के समय,
। दो समय तराजू तुलना कर रहे हैं, तो यह निष्कर्ष निकाला जा सकता है कि ताकना लिपिड प्रवाह करने के लिए थोड़ा प्रतिरोध प्रस्तुत करता है। हालांकि, अगर वास्तविक रिलीज के समय में काफी प्रसार सीमित रिहाई के लिए समय की तुलना में अब है, यह एक प्रक्रिया ऐसी ताकना चंचल के रूप में संकेत होता है, लिपिड रिहाई की गति को धीमा। प्रसार-सीमित रिलीज के समय,  , Fusing liposome और लिपिड diffusivity के आकार पर निर्भर करता है; अपने आकलन के इन दो मापदंडों मात्रा निर्धारित किया जा करने की आवश्यकता है। परख का एक अणु संवेदनशीलता लिपिड diffusivity हर संलयन घटना 26 के लिए एसबीएल में उनकी रिहाई के बाद कई एकल लिपिड fluorophores पर नज़र रखने से मापा जा करने की अनुमति देता है। हर fusing पुटिका के आकार(i) एक एकल लिपिड डाई की तीव्रता, (ii) एक डॉकिंग स्थल के आसपास की कुल प्रतिदीप्ति में परिवर्तन के बाद सभी fluorophores संलयन पर एसबीएल में स्थानांतरित कर रहे हैं, (iii) में जाना जाता लेबलिंग एसयूवी के घनत्व के संयोजन से 27 अनुमान लगाया जा सकता लिपिड, और (iv) लिपिड प्रति क्षेत्र। कई संलयन घटनाओं के लिए, वास्तविक लिपिड रिलीज के समय के रूप में पहले उल्लेख किया गया था वर्दी एसयूवी आकार 44 संभालने, प्रसार नियंत्रित रिहाई 27 तक होने की उम्मीद की तुलना में धीमी होना पाया गया है। मान लिया जाये कि लिपिड रिहाई की मंदता चंचल ताकना की वजह से है, एक मात्रात्मक मॉडल "ताकना खुलापन" के आकलन की अनुमति देता है, समय के अंश ताकना फ्यूजन 27 के दौरान खुला रहता है।
, Fusing liposome और लिपिड diffusivity के आकार पर निर्भर करता है; अपने आकलन के इन दो मापदंडों मात्रा निर्धारित किया जा करने की आवश्यकता है। परख का एक अणु संवेदनशीलता लिपिड diffusivity हर संलयन घटना 26 के लिए एसबीएल में उनकी रिहाई के बाद कई एकल लिपिड fluorophores पर नज़र रखने से मापा जा करने की अनुमति देता है। हर fusing पुटिका के आकार(i) एक एकल लिपिड डाई की तीव्रता, (ii) एक डॉकिंग स्थल के आसपास की कुल प्रतिदीप्ति में परिवर्तन के बाद सभी fluorophores संलयन पर एसबीएल में स्थानांतरित कर रहे हैं, (iii) में जाना जाता लेबलिंग एसयूवी के घनत्व के संयोजन से 27 अनुमान लगाया जा सकता लिपिड, और (iv) लिपिड प्रति क्षेत्र। कई संलयन घटनाओं के लिए, वास्तविक लिपिड रिलीज के समय के रूप में पहले उल्लेख किया गया था वर्दी एसयूवी आकार 44 संभालने, प्रसार नियंत्रित रिहाई 27 तक होने की उम्मीद की तुलना में धीमी होना पाया गया है। मान लिया जाये कि लिपिड रिहाई की मंदता चंचल ताकना की वजह से है, एक मात्रात्मक मॉडल "ताकना खुलापन" के आकलन की अनुमति देता है, समय के अंश ताकना फ्यूजन 27 के दौरान खुला रहता है।
जब भी व्यावहारिक है, यह दोनों लिपिड और घुलनशील सामग्री लेबल का उपयोग संलयन तंत्र का परीक्षण करने के लिए महत्वपूर्ण है। उदाहरण के लिए, लिपिड रिलीज में इस तरह के जाल प्रोटीन है कि टी चारों ओर से लिपिड प्रसार के प्रतिबंध के रूप में ताकना चंचल के अलावा अन्य प्रक्रियाओं, द्वारा मंद किया जा सकता हैवह ताकना। अगर इस मामले थे, तब, सामग्री की रिहाई लिपिड लेबल की रिहाई के पूर्व में होना होगा प्रदान की ताकना काफी बड़े घुलनशील जांच के पारित होने की अनुमति है। दृष्टिकोण में एक और अधिक मौलिक दोष धारणा एसबीएल लेबल लिपिड का हस्तांतरण एक पुटिका कि मोटे तौर पर अपनी पूर्व संलयन आकार को बरकरार रखा है एसबीएल जोड़ने के लिए एक संकीर्ण संलयन ताकना के माध्यम से होता है कि हो सकता है। एसबीएल में लिपिड हस्तांतरण भी, जैसा कि पहले लिपिड रिलीज डेटा अकेले 29 के आधार पर सुझाव दिया एक सहवर्ती साथ फ्यूजन ताकना, एसबीएल झिल्ली में एसयूवी की बेहद तेजी से पतन का तेजी से फैलाव से हो सकता है। दोनों लिपिड निगरानी और सामग्री को एक साथ रिलीज, यह पाया गया है कि कई pores अपने सभी लिपिड लेबल को जारी करने के बाद resealed, लेकिन उनके घुलनशील माल 27 के कुछ बरकरार रखा। यह इंगित करता है कि कम से कम कुछ liposomes संलयन के बाद एसबीएल में पतन नहीं, और एसबीएल में लिपिड डाई हस्तांतरण एक संलयन ताकना के माध्यम से होता है। इसके अलावा, एलipid और सामग्री जारी एक साथ 27 हुई, यह संभावना नहीं है कि लिपिड रिहाई की मंदता ताकना 45 के आस-पास जाल प्रोटीन द्वारा लिपिड प्रसार की बाधा के कारण किया गया था बना रही है।
एक एसयूवी-एसबीएल संलयन प्रोटोकॉल है कि घुलनशील सामग्री जारी की निगरानी नहीं था कि पहले Karatekin और रोथमान 25 द्वारा प्रकाशित किया गया था। इधर, अधिक हाल के घटनाक्रम शामिल किए गए हैं, लिपिड का अर्थात् एक साथ निगरानी और सामग्री जारी है और एसयूवी, लिपिड, और फ्यूजन ताकना गुण 27 के आकलन। प्रोटोकॉल एक गिलास coverslip 25 के साथ microfluidic कोशिकाओं, एक पाली (डाइमिथाइल siloxane) से संबंध बना (PDMS) elastomer ब्लॉक युक्त खांचे तैयार करने के लिए निर्देश के साथ शुरू होता है। फिर, दोनों लिपिड और सामग्री मार्कर के साथ वी-एसयूवी की तैयारी समझाया गया है। धारा 4 और 5, microfluidic कोशिकाओं कोडांतरण बगल में SBLs बनाने और दोषों और तरलता के लिए जाँच, वी.एस. की शुरूआत के लिए निर्देश प्रदानप्रवाह कोशिकाओं और फ्यूजन की घटनाओं का पता लगाने में यूवी। धारा 6 डेटा विश्लेषण के लिए निर्देश प्रदान करता है।
