Membranfusion är en universell biologisk process som krävs för intracellulär handel med lipider och proteiner, utsöndring, gödsling, utveckling och insvept virus inträde i värdorganismer 1-3. För de flesta intracellulära fusionsreaktioner inklusive frisättning av hormoner och signalsubstanser via exocytos, är energin för att sammansmälta två lipiddubbelskikt tillhandahålls genom bildning av ett fyra-helix bundle mellan cognate löslig N-etylmaleimid-känslig faktor fastsättning proteinreceptor (SNARE) proteiner, förankrad i vesikeln (v-SNARE) och målmembranet (t-SNARE) 4, respektive. Synaptic vesikler exocytos är den mest hårt reglerad fusion reaktion och inträffar inom en millisekund efter ankomsten av en aktionspotential 1,4,5. Fusions por, den första anslutningen mellan de två fixerings facken, kan flimmer öppna och slutna flera gånger innan återförslutning eller expandera irreversibelt 5-7. De tidigare resultatövergående, "Kiss & run" fusion, medan den senare leder till full fusion. Faktorer som styr balansen mellan dessa två typer av fusion och mekanismer som reglerar por flimmer är inte väl förstått 5,8.
SNARE proteiner krävs för exocytos; synaptiska vesikelfusion avskaffas vid klyvning av snaror av neurotoxiner 9. Bulkfusionsexperiment med användning av små unilamellära vesiklar (SUVs) visade att snaror inte bara krävs, men också tillräcklig för att driva membranfusion 10. I detta bulk assay, SUVs rekonstitueras med v-snaror (v-SUV) till dopad med fluorescerande fosfolipider (N – (7-nitro-2-1,3-bensoxadiazol-4-yl) -phosphoethanolamine (NBD-PE) och ( N -. (lissaminrodamin B sulfonyl) -phosphoethanolamine (LR-PE) och blandades med omärkta vesiklar innehållande t-snaror (t-SUV) Initialt fluorescensen av NBD-PE i v-SUVs släckes genom Förster resonansenergiöverföring (FRET ) till LR-PE. som labELED v-SUV smälta samman med omärkta t-SUV, är fluoroforen ytdensitet i det nu kombinerade membranet reduceras och den resulterande ökningen i NBF-PE fluorescens rapporterar omfattningen av lipidblandning 10. Som bulk analysen är lätt att ställa upp och analysera, har det i stor utsträckning använts för att studera mekanismerna för SNARE-medierad sammansmältning 10-14. Det har dock flera begränsningar, såsom låg känslighet och dålig tidsupplösning. Viktigast som en ensemble mätning, det genomsnitt resultat under alla omständigheter gör diskriminering mellan dockning och fusion, samt detektering av hemifusion mellan svårt.
Under det senaste årtiondet flera grupper, inklusive vårt, har utvecklat nya analyser för att övervaka fusions evenemang på enstaka vesikler nivå 15-27. Ha och kollegor använde v-SUV bundna på en yta och övervakas deras fusion med gratis t-SUV 18,19. Lipidblandning övervakades med användning av FRET mellan ett par lipid-bundna fluoroforer embäddas in i v- och t-SUV, respektive, med hjälp av total inre reflektion fluorescens (TIRF) mikroskopi 18. Senare Brunger laboratorium använde en enda lipid-label art tillsammans med en innehåll markör för samtidig detektion av lipid och innehåll blanda 20,28. Både lipid och innehållet markörerna ingick vid höga, självläckande koncentrationer; fusion med omärkta SUVs resulterade i fluorescens dequenching 20,28.
Andra har smält v-SUV till plana bilayers ombildade med t-snaror 15-17,21-27,29. Den plana geometri målet (t-SNARE innehållande) biskikt bättre efterliknar den fysiologiska fusionsprocessen av små, mycket böjda vesiklar med platt plasmamembranet. Den Steinem grupp som användes pore överbryggande membran rekonstituerade med t-snaror, suspenderade genom ett poröst kiselnitrid substratet och detekteras fusion med enskilda v-SUVs med användning av konfokal laserscanningsmikroskopi 23. andra fbegagnade v-SUVs till plana biskikt rekonstituerade med t-snaror, uppburen på ett glassubstrat 15-17,21,22,24-27,29. Den stora fördelen med att använda stöds dubbla skikt (SBLs) är att TIRF mikroskopi kan användas för att detektera dockning och fusion händelser med utmärkt signal-brusförhållande och utan inblandning från fria v-SUV, även om användning av mikrofluidik ger också enstaka händelse upplösning med standard fjärrfälts epifluorescensmikroskopi 24.
En viktig fråga är om och hur substrat tvåskiktsmembran interaktioner påverkar stöds tvåskiktsmembran kvalitet och fusionsprocessen. Tidigt arbete utnyttjade plana SBLs som direkt stöd på ett glas eller kvartssubstrat 15-17. Dessa SBLs gjordes genom adsorption, spricker, spridning och blandning av t-SUV membran på substratet. Det var snart insåg dock att utelämna en viktig t-SNARE komponent, SNAP25 från SBLs framställda på detta sätt resulterade i v-SUV dockning och fusion kinetik indistinguisHable från de som erhållits genom att använda den fullständiga t-snaror 17. Eftersom SNAP25 är absolut nödvändig för fusion in vivo 30,31, var den fysiologiska relevansen av dessa tidiga försök ifrågasättas. Tamm grupp klarade denna utmaning genom att använda kontrolleras bättre stöd tvåskiktsmembran bildning 21. Det brukade Langmuir-Blodgett avsättning för proteinfritt första broschyr av SBL, följt av fusion av det monolager med t-SUV 21. Detta resulterade i SNAP25 beroende fusion.
För att undvika potentiella artefakter associerade med ett dubbelskikt direkt uppburen på ett glassubstrat utan behov av att använda Langmuir-Blodgett metoder, Karatekin et al. Infört en mjuk, hydratiserad poly (etylenglykol) (PEG) kudde mellan dubbelskiktet och substratet 24. Denna modifiering resulterade också i SNAP25 beroende fusion 24. Dubbelskikt vadderade på ett mjukt polymerskikt hade varit kända för att bättre bevara transprotein rörlighet och funktion 32, och hade använts i fusionsstudier med virus 33. Dessutom PEGylerade bilayers tycks behålla en del förmåga att självläka och är mycket robust 34,35. Först är en fraktion av kommersiellt tillgängliga, lipid-länkad PEG-kedjor som ingår i t-SUV membran. När dessa t-SUVs brista och bilda en plan tvåskiktsmembran på ett glassubstrat, omfattar en PEG borste båda broschyrer av den plana biskikt. Eftersom plana biskikt bildning drivs av vidhäftning av PEG-kedjor närliggande t-SUVs på den hydrofila glasytan, liposom sprängning och plana biskikt bildning är relativt okänsliga för lipidkompositionen användas. När emellertid stora mängder kolesterol ingår, ökar de kohesiva egenskaperna hos SUVs, SUVs kanske inte brista spontant. Om detta är fallet, kan osmotisk chock eller tvåvärda joner användas för att hjälpa till plana biskikt bildning 25.
Såsom nämnts ovan, i detta apgrepp en PEG borste täcker båda sidorna av den plana, uppburen dubbelskiktet. Borsten mot mikroflödesflödeskanalen hjälper till att förhindra ospecifik vidhäftning av inkommande v-SUV som också är oftast täckt med en PEG lager. Bildning av V- och t-SNARE-komplex utgår från membran distala N-terminaler och fortsätter i etapper mot membran-proximala domänerna 36. För v-SUV att interagera med t-SBL, V- och t-SNARE N-terminaler är nödvändigt att skjuta över PEG borstar, som verkar vara fallet i enlighet med villkoren i analysen. Borsthöjden kan anpassas för att studera andra än snaror proteiner genom att variera densiteten hos PEGylerade lipider och PEG kedjelängd 37,38. En annan fördel med de PEG-borstar som täcker de proximala ytorna av fixerings bilayers är att de efterliknar det trånga miljön i biologiska membran, vilka är packade med 30,000-40,000 integrerade membranproteiner per kvadratmikron 39. Precis som de PEG-kedjor i denna analys, den Repulsive proteinskikt som täcker biologiska membran måste skjutas åt sidan för att tillåta kontakt mellan de två fosfolipid bilayers för fusion skall ske.
Mikroflödesströmningskanaler används i denna analys, eftersom de erbjuder unika fördelar. Först gör mikroflödes flöde jämnare avsättning av T-SUV för att sprida och säkring för att bilda t-SBL. För det andra, (<1 l) minimerar den lilla kanalvolymen provförbrukning. För det tredje, de små volymer som behövs låta hela experimentet genomföras under konstant flöde. Flow avlägsnar svagt, förmodligen ospecifikt, vidhäftade v-SUV från SBL 16. Det har även en konstant densitet av v-SUV ovanför t-SBL, förenklar kinetisk analys 17. Slutligen dockade blåsor lätt skiljas från fri dem som bärs av flödet 25. Fjärde flera mikroflödessystem kanaler kan användas på samma täck varje sondering olika tillstånd. Detta möjliggör jämförelser av villkor During samma experimentkörning. Ett liknande tillvägagångssätt har använts av van Oijen för att studera fusion mellan influensavirus och vadde SBLs 33.
I TIRF mikroskopi, den exponentiella avklingningen av det evanescenta fältet (med en sönderfallskonstant ~ 100 nm) begränsar fluorescensexcitation till de molekyler som är i mycket nära anslutning för det glasbuffertgränssnittet. Detta minimerar bidraget från fluorescerande molekyler som ligger längre bort, ökar signal-brusförhållande, och tillåter enda molekyl känslighet med ram exponeringstider på 10 till 40 msek. Det evanescenta fältet leder också till en signalökning på fusion: som den märkta lipider överföring från SUV till SBL, de befinner sig, i genomsnitt, en starkare exciteringsfältet. Denna ökning i fluorescens är starkare för större liposomer.
Om polariserat ljus används för att generera det evanescenta fältet, ytterligare effekter bidra till förändringar i fluorescens vid tverför av etiketter från SUV i SBL. Vissa lipid-färgämnen har en övergångs dipol orienterad med en föredragen medel vinkel med avseende på dubbelskiktet i vilket de är inbäddade. Detta skapar en skillnad i mängden av fluorescens som emitteras av de fluoroforer när de är i SUV kontra SBL, eftersom den polariserade strålen att excitera färgämnen i de två membranen på olika sätt. För de förstnämnda kommer excitationsstrålen interagera med övergångs dipoler orienterade kring den sfäriska SUV, medan det senare kommer dipol riktlinjer begränsas av den plana SBL geometri. Till exempel, när s-polariserade infallande ljuset (polariserat vinkelrätt mot infallsplanet) används, är excitering mer effektiv när färgämnet är i SBL än i SUV för en lipid färgämne övergångs dipol orienterad parallellt med membranet 29,40 (såsom den av Dil eller gjorde 41-43). En SUV dopad med en sådan fluorofor visas dim när den dockar har laddats in i SBL (Figur 7, Representat ive Resultat). Som en fusion pore öppnas och förbinder de SUV och SBL membran, fluorescerande prober diffundera in i SBL och blir mer sannolikt att exciteras av s-polariserade evanescent fält 25,27,29. Följaktligen ökar fluorescenssignalen integreras runt fusionsstället kraftigt under färgöverföring från SUV till SBL 27 (Figur 3 och Figur 7). En ytterligare faktor som bidrar till signal förändringar som följer fusion dequenching av fluorescerande markörer som de späds när de överförs in i SBL. Bidraget från dequenching är vanligtvis små i jämförelse med evanescent fältröta och polariseringseffekter i analysen beskriven här, eftersom endast en liten fraktion ( 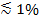 ) Av lipiderna är märkta.
) Av lipiderna är märkta.
Signalökning vid fusion kan utnyttjas för att härleda fusion pore egenskaper genom att jämföra den tid,1 "src =" / filer / ftp_upload / 54349 / 54349eq2.jpg "/>, som krävs för en lipid fly genom en por som är fritt permeabel för lipider till frigöra tid själva,  . Om de två tidsskalor är jämförbara, skulle man dra slutsatsen att porerna uppvisar litet motstånd mot lipid flödet. Men om frigöra tid själva är betydligt längre än den tid för diffusion begränsad frisättning, skulle detta tyda på en process, såsom porer flimmer, fördröjer lipid release. Spridning begränsade frigöra tid,
. Om de två tidsskalor är jämförbara, skulle man dra slutsatsen att porerna uppvisar litet motstånd mot lipid flödet. Men om frigöra tid själva är betydligt längre än den tid för diffusion begränsad frisättning, skulle detta tyda på en process, såsom porer flimmer, fördröjer lipid release. Spridning begränsade frigöra tid, 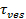 , Beror på storleken av fixerings liposomer och lipid-diffusivitet; sin uppskattning kräver dessa två parametrar som skall kvantifieras. Den enda molekyl känsligheten för analysen tillåter lipid diffusivitet som skall mätas genom att spåra flera enda lipid fluoroforer efter frigivningen i SBL för varje fusion händelse 26. Storleken på varje fusing vesikelkan uppskattas 27 genom att kombinera (i) intensiteten av en enda lipid färgämne, (ii) förändring av det totala fluorescens runt en dockningsområdet efter det alla fluoroforer överförs till SBL upon fusion, (iii) den kända märkningsdensitet av SUV lipider, och (iv) området per lipid. För många fusion händelser, var de faktiska lipid frisättningstider visat sig vara mycket långsammare än väntat genom diffusion-kontrollerad frisättning 27, som noterades tidigare antagande enhetlig SUV storlek 44. Förutsatt fördröjning av lipid frisättning beror på pore flimmer, gör en kvantitativ modell uppskattning av "pore öppenhet", den del av tiden porerna förblir öppen under fusion 27.
, Beror på storleken av fixerings liposomer och lipid-diffusivitet; sin uppskattning kräver dessa två parametrar som skall kvantifieras. Den enda molekyl känsligheten för analysen tillåter lipid diffusivitet som skall mätas genom att spåra flera enda lipid fluoroforer efter frigivningen i SBL för varje fusion händelse 26. Storleken på varje fusing vesikelkan uppskattas 27 genom att kombinera (i) intensiteten av en enda lipid färgämne, (ii) förändring av det totala fluorescens runt en dockningsområdet efter det alla fluoroforer överförs till SBL upon fusion, (iii) den kända märkningsdensitet av SUV lipider, och (iv) området per lipid. För många fusion händelser, var de faktiska lipid frisättningstider visat sig vara mycket långsammare än väntat genom diffusion-kontrollerad frisättning 27, som noterades tidigare antagande enhetlig SUV storlek 44. Förutsatt fördröjning av lipid frisättning beror på pore flimmer, gör en kvantitativ modell uppskattning av "pore öppenhet", den del av tiden porerna förblir öppen under fusion 27.
Närhelst praktiskt, är det viktigt att testa fusionsmekanismer med användning av både lipid- och lösliga innehåll etiketter. Till exempel kan lipid frisättning fördröjas av andra än por flimmer processer, såsom begränsning av lipid diffusion av snaran proteiner som omger than pore. Om så vore fallet, släpp sedan av innehållet skulle föregå frigörande av lipid etiketter, förutsatt att por är tillräckligt stor för att tillåta passage av lösliga sonder. En mer grundläggande brist i den strategi skulle kunna vara i antagandet att överföringen av märkta lipider till SBL sker genom en smal fusion por ansluter SBL till en vesikel som till stor del har behållit sin pre-fusion form. Lipid överföring till SBL kan också vara resultatet av en snabb utvidgning av fusions por med en åtföljande, extremt snabb kollaps av SUV i SBL membran, som föreslog tidigare på lipid släppuppgifter enbart 29. Övervakning både lipid och innehåll släpper samtidigt visade det sig att många porer återförslutas efter att ha släppt alla sina fettetiketter, men behöll en del av deras lösliga last 27. Detta tyder på att åtminstone vissa liposomer inte kollapsar in i SBL efter smältning, och att lipiden färgämnesöverföring in i SBL sker genom en fusion por. Dessutom, lIPID och innehåll frisättning inträffade samtidigt 27, vilket gör det osannolikt att fördröjning av lipid frigör berodde på besvär av lipid diffusion av snaran proteiner omger por 45.
En SUV-SBL fusionsprotokoll som inte övervaka lösliga innehåll frisättning har tidigare publicerats av Karatekin och Rothman 25. Här är den senaste utvecklingen ingår, nämligen samtidig övervakning av lipid och innehåll släpper och uppskattning av SUV, lipid, och fusion pore egenskaper 27. Protokollet börjar med instruktioner för framställning av de mikroflödesceller, tillverkade genom bindning av en poly (dimetylsiloxan) (PDMS) elastomer segmentinnehållande räfflor med ett täckglas 25. Därefter framställning av v-SUV med både lipid och innehåll markörer förklaras. §§ 4 och 5 ger instruktioner för montering av mikroflödesceller, bildar SBLs in situ och kontroll av defekter och smidighet, införande VSUVS in flödescellerna och detektering av fusionshändelser. Avsnitt 6 innehåller instruktioner för dataanalys.
