analisi della crescita dipende da una serie di strumenti che vengono comunemente utilizzati dagli scienziati vegetali per descrivere genotipo determinato differenze di crescita e / o risposte fenotipiche a fattori ambientali. Essi comprendono dimensione e peso misurazioni di tutta la pianta o di un organo e calcoli dei tassi di crescita per esplorare i meccanismi alla base della crescita. Crescita degli organi è determinata dalla divisione cellulare e l'espansione a livello cellulare. Pertanto, compresa la quantificazione di questi due processi in crescita analizza è la chiave per comprendere le differenze nella crescita dell'intero organo-1. Pertanto, è fondamentale disporre di un metodo adeguato per determinare i parametri di crescita cellulare che è relativamente facile da usare da laboratori non specializzati.
Analisi cinematica è già stato stabilito come un approccio che fornisce un quadro potente per lo sviluppo di modelli di crescita di organi 2. La tecnica è stata ottimizzata per sistemi lineari,come radici Arabidopsis thaliana e foglie monocotiledoni, ma anche per i sistemi non lineari, come foglie dicotiledoni 3. Al giorno d'oggi, questa metodologia è sempre più utilizzato per studiare come genetici, ormonali, dello sviluppo e fattori ambientali influenzano la divisione cellulare e di espansione in vari organi (Tabella 1). Inoltre, fornisce anche un quadro di collegare processi cellulari ai loro regolamenti biochimici, molecolari e fisiologici sottostanti (Tabella 2), anche se le limitazioni possono essere imposte in base alle dimensioni degli organi e l'organizzazione spaziale per tecniche che richiedono una maggiore quantità di materiale vegetale (ad esempio, di metaboliti misurazioni, proteomica, etc.).
Foglie monocotyledonous, come il mais (Zea mays) foglie, rappresentano sistemi lineari in cui le cellule si spostano dalla base della foglia verso la punta, sequenzialmente passando attraverso il meristema e l'allungamento zona per raggiungere la maturitàzona. Questo lo rende un sistema modello ideale per studi quantitativi dei modelli spaziali di crescita del 4. Inoltre, foglie di mais hanno zone di forte crescita (meristema e zona di allungamento che abbracciano diversi centimetri 5) e forniscono possibilità per gli studi ad altri livelli organizzativi. Questo permette la ricerca dei (presunti) meccanismi regolatori che controllano la divisione cellulare e di espansione, quantificato da analisi cinematica attraverso una serie di tecniche molecolari, misurazioni fisiologiche, e approcci di biologia cellulare (Tabella 2).
Qui, forniamo un protocollo per l'esecuzione di una analisi cinematica in foglie monocot. In primo luogo, spieghiamo come condurre una corretta analisi sia di divisione cellulare e l'allungamento delle cellule in funzione della posizione lungo l'asse foglia e come calcolare i parametri cinematici. In secondo luogo, mostriamo anche come questo può essere utilizzato come base per il disegno di campionamento. Qui, discutiamo due casi: ad alta risoluzione di campionamento di und focalizzata campionamento, consentendo una migliore interpretazione dei dati e il risparmio di tempo / denaro, rispettivamente.
Tabella 1. Panoramica di cinematica analizza i metodi per la quantificazione della divisione cellulare e di espansione in vari organi.
| organo | riferimento |
| foglie monocotiledoni | 16, 20, 21, 22 |
| apici radicali | 2, 23, 24, 25, 26, 27, 28, 29 |
| foglie dicotiledoni | 21, 30, 31 |
| sparare meristema apicale | 32 |
Tabella 1. Panoramica di cinematica analizza i metodi per la quantificazione della divisione cellulare e di espansione in vari organi.
<p class="jove_content" fo:keep-together.within-page = "1">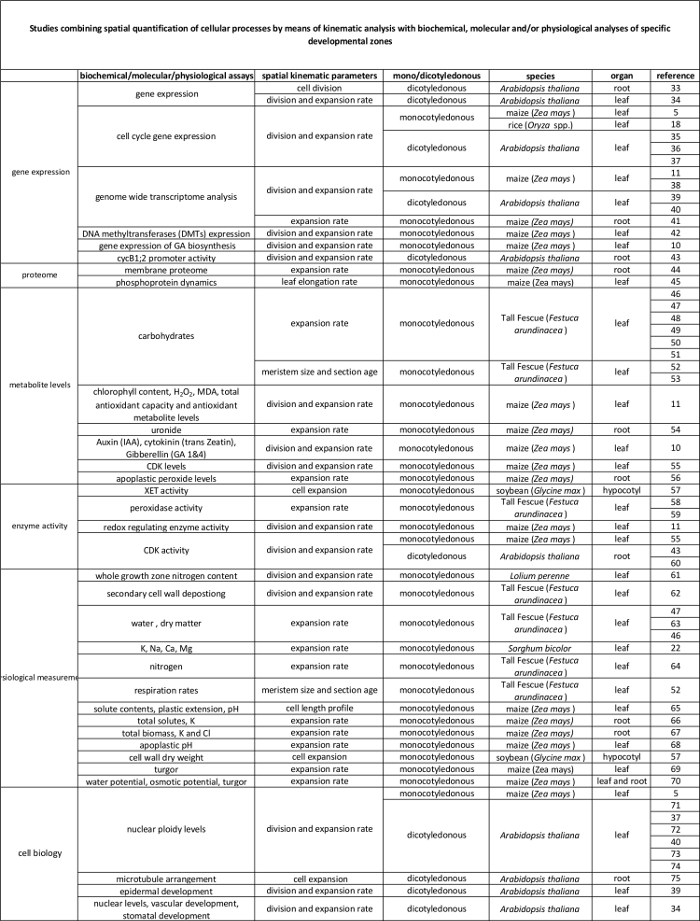
Tabella 2. Collegamento tra processi cellulari quantificati dall'analisi cinematica per la loro regolamentazione a livello molecolare. I riferimenti ai vari studi che collegano la quantificazione dei processi cellulari ai risultati di saggi biochimici e molecolari di varie specie e organi. Endotransglucosylase Xyloglucan (XET), malondialdeide (MDA), chinasi ciclina-dipendenti (CDK). Clicca qui per vedere una versione più grande di questa tabella.
