Análise de crescimento depende de um conjunto de ferramentas que são comumente usados por cientistas de planta para descrever as diferenças de crescimento genótipo determinado e / ou respostas fenotípicas a fatores ambientais. Eles incluem tamanho e peso medições da planta inteira ou um órgão e cálculos de taxas de crescimento para explorar os mecanismos subjacentes de crescimento. Crescimento dos órgãos é determinada por divisão celular e de expansão ao nível celular. Portanto, incluindo a quantificação desses dois processos no crescimento análises é fundamental para compreender as diferenças no crescimento do órgão inteiro 1. Por conseguinte, é fundamental ter uma metodologia adequada para determinar os parâmetros de crescimento celular que é relativamente fácil de usar por laboratórios não especializados.
A análise cinemática já foi estabelecida como uma abordagem fornecendo uma estrutura poderosa para o desenvolvimento de modelos de crescimento de órgãos 2. A técnica foi optimizado para sistemas lineares,tais como raízes e folhas de Arabidopsis thaliana monocotiledóneas, mas também para os sistemas não lineares, tais como folhas dicotiledóneas 3. Hoje em dia, esta metodologia está cada vez mais sendo usado para estudar como genética, hormonal, de desenvolvimento e fatores ambientais influenciam a divisão celular e expansão em vários órgãos (Tabela 1). Além disso, também fornece uma estrutura para conectar processos celulares aos seus regulamentos bioquímicas, moleculares e fisiológicos subjacentes (Tabela 2), embora as limitações podem ser impostas por tamanho do órgão e da organização espacial para técnicas que exigem maiores quantidades de material vegetal (por exemplo, metabólitos medições, proteômica, etc.).
Folhas de monocotiledóneas, como o milho (Zea mays) da folha, representam sistemas lineares em que as células se movem a partir da base da folha em direcção a ponta, passando sequencialmente através da zona de meristema e alongamento para atingir a maturidadezona. Isto o torna um sistema modelo ideal para estudos quantitativos dos padrões espaciais de crescimento de 4. Além disso, folhas de milho têm zonas de grande crescimento (meristema e zona de alongamento abrangendo vários centímetros 5) e proporcionar possibilidades para estudos em outros níveis organizacionais. Isto permite a investigação dos mecanismos de regulação (putativos) que controlam a divisão celular e expansão, quantificada por análise cinemática através de uma série de técnicas moleculares, medições fisiológicas e abordagens de biologia celular (Tabela 2).
Aqui, nós fornecemos um protocolo para a realização de uma análise cinemática em folhas de monocotiledôneas. Em primeiro lugar, vamos explicar como conduzir uma análise adequada de ambos divisão celular e alongamento celular como uma função da posição ao longo do eixo da folha e como calcular parâmetros cinemáticos. Em segundo lugar, também mostrar como este pode ser usado como uma base para a concepção de amostragem. Aqui, discutimos dois casos: de alta resolução de uma amostragemd focado amostragem, permitindo uma melhor interpretação dos dados e economia de tempo / dinheiro, respectivamente.
Tabela 1. Visão Geral de cinemática analisa métodos de quantificação da divisão celular e expansão em vários órgãos.
| órgão | referência |
| folhas monocotiledôneas | 16, 20, 21, 22 |
| dicas de raiz | 2, 23, 24, 25, 26, 27, 28, 29 |
| folhas de dicotiledôneas | 21, 30, 31 |
| atirar meristema apical | 32 |
Tabela 1. Visão Geral de cinemática analisa métodos de quantificação da divisão celular e expansão em vários órgãos.
<p class="jove_content" fo:keep-together.within-page = "1">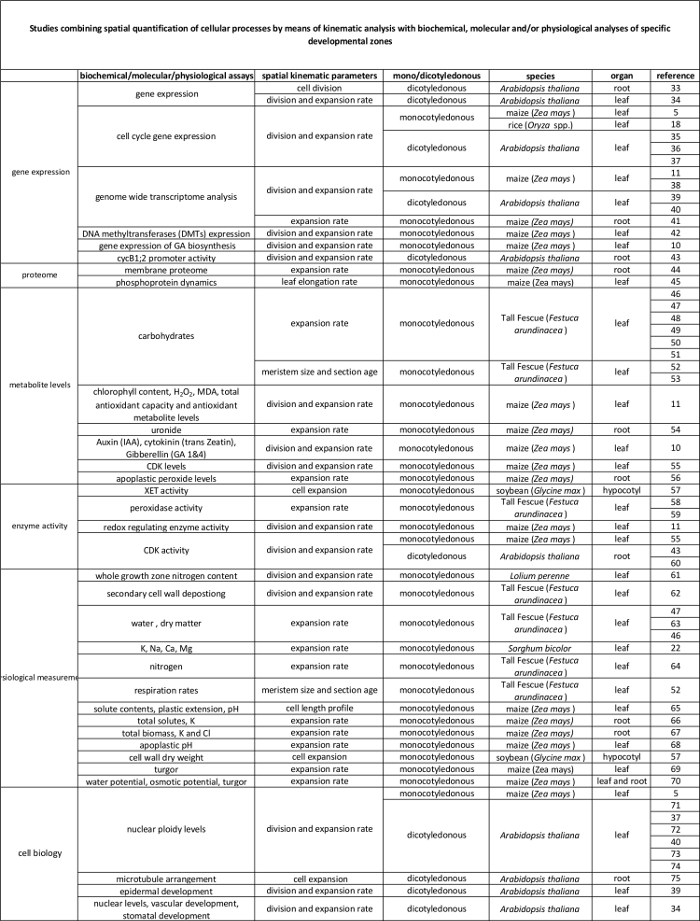
Tabela 2. A ligação entre os processos celulares quantificados pela análise cinemática a sua regulação ao nível molecular. As referências a diversos estudos de ligação a quantificação de processos celulares com os resultados de ensaios bioquímicos e moleculares em várias espécies e órgãos. Endotransglucosylase xiloglucano (XET), malondialdeído (MDA), quinases dependentes da ciclina (CDK). Por favor clique aqui para ver uma versão maior desta tabela.
